College of Social and Behavioral Sciences
113 Influence of Nature on Effortful Processing of Emotionally Charged Stimuli
Astra Storie and Amy McDonnell
Faculty Mentor: Amy S. McDonnell (Psychology, University of Utah)
Abstract
Immersion in natural environments is thought to have a restful and restorative effect on individuals living in urban areas. Attention Restoration Theory (ART) suggests that depletion of attentional resources strained by life in urban environments is a primary driver of the restorative benefits of nature, and is substantiated by a body of evidence showing the benefits of nature on cognitive processes related to attention. This study sought to examine if immersion in nature benefits two aspects of attention– cognitive control and emotional inhibition–as assessed by a Classic Stroop Task and Emotional Stroop Task, respectively. Participants (N=27) completed self-report measures and both cognitive tasks once in an urban environment, and once in a natural environment during a five-day camping trip. We predicted that performance on the Classic Stroop Task and on the Emotional Stroop Task would improve in nature compared to the urban control environment. Contrary to our hypotheses, linear mixed models did not show a significant impact of environment type on performance on either task. We did, however, find that self-reported connectedness to nature increased in natural environments. These results suggest no effect of immersion in nature on behavioral indices of cognitive control or emotional inhibition, but did show that immersion in nature enhances perceived connectedness to nature.
Introduction
Urbanization has rapidly increased since the industrial revolution of the 19th century, resulting in 56% of the global population living in urban environments (The World Bank, 2023). Epidemiological research has found that populations living in urban environments experience greater rates of mental illness (Golembiewski, 2021) and decreased attentional capacity (Linnell et al., 2012; Lambert et al., 2015). Although the restorative properties of natural environments have been known since antiquity, it has only been in the last several decades that these effects have been tested with the scientific method by the psychological science community. This research has resulted in a growing body of research in support of the restorative properties of nature on cognition and mood.
One framework that explains how these benefits may precipitate from interacting with nature is Attention Restoration Theory (ART; Kaplan, 1995). ART suggests that the over- stimulation and attentional demands of urban environments place high demand on our voluntary, directed attention. We are forced to move from one task to the next and we are unable to fully replenish our cognitive resources before this effortful form of attention is next required. Over time, this constant task-switching leads to mental fatigue, reduction in our voluntary attention capacity, degraded cognitive performance, and increased stress. Natural environments, by contrast, feature stimuli which are inherently pleasing or salient such as vast landscapes, wildlife, and diverse vegetation. These features engage our more passive, involuntary attention in a less effortful manner. This reduction in use of directed attention results in the restoration of attentional resources, and subsequent improvements to cognitive performance and mood (Kaplan, 1995).
ART is supported by research that shows exposure to nature benefits multiple facets of attention including cognitive flexibility (Hartig, 1991), response inhibition (Bailey et al., 2018), creativity (Atchley et al., 2012), and working memory capacity (Berman et al., 2012).Additionally, exposure to nature has been shown to improve affect (Berman et al., 2012; Bratman et al., 2015), anxiety, and stress (Koselka et al., 2019). Although ART suggests that nature primarily restores attention and the emotional benefits are dependent on this restoration, contemporary research points toward these benefits being interrelated (Scott et al., 2021). For example, self-reported connectedness to nature has even been shown to partially mediate the positive effects of nature on cognitive processes (Mayer et al., 2008).Additionally, recent research has demonstrated that children tested in more natural “green” spaces exhibit less distraction from negative emotional stimuli than those tested in indoor settings (Mason et al., 2022). These results are particularly important as recent research has found cognitive control to have a limited ability to improve emotional distraction (Straub et al., 2021). It is more likely that cognitive control and emotional distraction are related, and draw from shared attentional resources instead of being dependent on the restoration of attention alone.
A common means of assessing cognitive control is the use of Stroop tasks (Stroop, 1935), which require participants to exert effortful, inhibitory control to sort presented stimuli. Variations exist such as the Emotional Stroop task which specifically engages emotional response inhibition. Since these tasks require directed, effortful attention they have been used in previous studies to demonstrate the restorative effect of nature on attention (Bailey et al., 2018). Benefits to both a Classic and Emotional Stroop task after exposure to nature would support the emerging view that cognitive control (Classic Stroop Task) and emotional cognitive control (Emotional Stroop Task) operate using shared resources. Such findings would also implicate that the restorative effect of nature extends beyond directed attention to benefit other processes such as emotional control.
The present study seeks to investigate the benefits of immersion in nature on cognitive control and emotional response inhibition–two aspects of higher-order executive attention. We assessed behavioral performance on the Classic Stroop Task and the Emotional Stroop Task while participants were immersed in the wilderness on a five- day camping trip compared against control testing in an urban environment. We examined if immersion in nature impacts cognitive inhibitory control, emotional inhibitory control, or both. Furthermore, we examined if self-reported connectedness to nature strengthens or weakens the impact of immersion in nature on these cognitive processes, as observed in previous studies (Mayer et al., 2008). Improvement on both tasks would suggest that immersion in nature enhances cognitive control capacity not just for neutral stimuli but also emotionally charged stimuli, thus suggesting the attentional benefits of exposure to nature are domain-general.
Methods
Participants
Twenty-seven undergraduate students (15 females, 11 males, and 1 other; Mage = 22.63; SD = 3.80) at the University of Utah with normal neurological functioning, normal or corrected-to-normal vision, and fluent in English participated in the study as part of a course at the University of Utah. While attendance on the nature trip was required for course credit, participation in the presented research study was voluntary and participants were paid $60 for completion of the research study.
Design
Each participant completed testing at two time points–once in a natural environment and once in an urban environment– for a within-subjects design. The order of testing was counterbalanced across participants to control for potential practice effects.
Measures
The Stroop task (Stroop, 1935) is a widely used cognitive test to assess a facet of attention called inhibitory cognitive control. The task presents participants with the names of colors (red, blue, yellow, or green) in different colored fonts (red, blue, yellow, or green) and requires the participant to respond to the color of the font while ignoring lexical meaning. For example, if the word “red” was presented in blue ink, the correct answer would be “blue”. The difference in reaction times where the lexical meaning and font color are incongruent and the trials where they are congruent (i.e., response to “RED” printed in blue ink minus “RED” printed in red) is referred to as the Stroop Effect. Smaller Stroop Effect scores indicate greater inhibitory control in participants. The Emotional Stroop task (Ben-Haim et al., 2016) is similar to the original Stroop task in that participants are required to respond to the font color while ignoring lexical meaning. In the Emotional Stroop task however, the presented words are common English words which either have a neutral valence (e.g., “FIELD”, “NEAR”, “BLOOM”), or an emotional valence (e.g., “DEATH”, “FAILURE”, “GRIEF”). The difference between the response times to the emotionally charged trials and the neutral trials is referred to as the Emotional Stroop Effect. Smaller Emotional Stroop Effect scores indicate reduced interference from the emotional valence of the word and indicate less emotional distraction in participants.
Connectedness to nature was assessed with the Connectedness to Nature scale (Mayer & Frantz, 2004), which has previously been shown to partially mediate positive emotional and cognitive benefits to exposure to nature (Mayer et al., 2009). This inventory consists of fourteen questions assessing affective responses of an individual in response to nature. Responses are recorded on a 5-point scale where 1 indicates the least connection with nature, and 5 the highest. After adjusting for reverse-coded items, total scores are calculated as an average of the 14 questions on the scale.
Other Measures
Other self-report measures and cognitive tasks were administered as part of the original study to address other research questions not pertinent to the present analysis. They are included in the procedure for clarity and comprehensiveness, but will not be discussed in the present analysis.
Procedure
After participants provided informed consent, they were randomly assigned to one of two groups. Participants completed testing either before and during (Group 1) or during and after (Group 2) a five-day camping trip at Sand Island Campground in Bluff, UT. This counterbalanced AB-BA design was chosen to allow for a testing session during the immersion in nature and a control, urban testing session for each participant, while controlling for practice effects from repeated measures. The urban testing session of Group 1 was conducted one week prior to the trip, while the urban testing session for Group 2 was conducted one week after the camping trip.
The nature testing sessions for all participants occurred on either day three or four of the camping trip, outside of time reserved for meals and class related activities. Testing occurred at a campsite approximately 200 yards away from the class campsite in order to separate research activities from class activities, and to minimize distractions during testing. During testing, participants were seated in a foldable camp chair inside of a pop-up nylon shelter as shown in Figure 1 to protect from glare on the testing screen and natural elements such as wind, rain, and insects. Participants were sat in the same conditions in the urban condition for testing consistency. The remainder of the study procedure was identical for all testing sessions, only differing in location of testing.

Figure 1: Example testing setups in the urban condition (left) and natural condition (right). At the beginning of a testing session, participants completed an IRB-approved consent document and general demographic survey to verify they met the study requirements.
Participants then completed the Operation Span test, Stroop Task, and then Emotional Stroop Task on a laptop provided by the experimenters. Written instructions and a brief practice test were given for each cognitive task, and a break of 10 minutes was given between these tasks. Participants then took the Reading The Mind In The Eyes task on paper. Finally participants responded to the Perceived Positive and Negative Affect Scale (PANAS), Connectedness to Nature Scale (CNS), a social connection scale, sleep scale, and physical activity scale.
Analysis
Statistical analyses were conducted using R software, version 4.3.1. On both the classic Stroop and the emotional Stroop task, reaction times that were 3 standard deviations above the mean response time, or that were under 100 ms were considered to be the result of inattention to the task and were removed from future analyses. For the classic Stroop task, each participant’s mean response time for congruent trials was subtracted from their mean response time to incongruent trials in each testing environment. Greater differences between these reaction time means indicated greater interference from incongruence of the ink and lexical color of the stimuli, and indicates decreased cognitive control capacity. Similarly, for the Emotional Stroop task, each participant’s mean response time for neutral trials was subtracted from their mean response time to emotional trials. Greater differences indicated greater interference from the negative valence of the word, and indicated decreased cognitive control in suppressing emotional responses. The Perceived Connectedness to Nature responses were converted to scores of 1-4 for each question, and averaged into one total score for each participant at each testing session.
To account for anticipated variability between and within groups, repeated measures within each individual, and missing data, data were analyzed using linear mixed effects models (Bates et al., 2015). For each outcome measure of interest, three models were run such that Environment Type, Session, and the interaction between Environment Type and Session were each entered one at a time as fixed effect predictors with Subject ID as the random intercept. A subsequent likelihood ratio test was run to test for significance of each model in which the model of interest was compared to a null model with the variable of interest removed. This test results in a chi-squared statistic and associated p-value for each model. Effect sizes for significant effects are reported as Cohen’s d scores.
Results Self-report
Results
The results from the self-report Connectedness to Nature Scale can be visualized in Figure 2. Linear mixed models revealed a significant main effect of environment type on score on the connectedness to nature scale (χ2(1)=7.16, p=0.00745) such that there was a significant increase in the connectedness to nature scale score in nature compared to urban environments (β = 0.21, SE=0.076, df=26.00, t=2.81, p=0.00928; Cohen’s d=0.38)
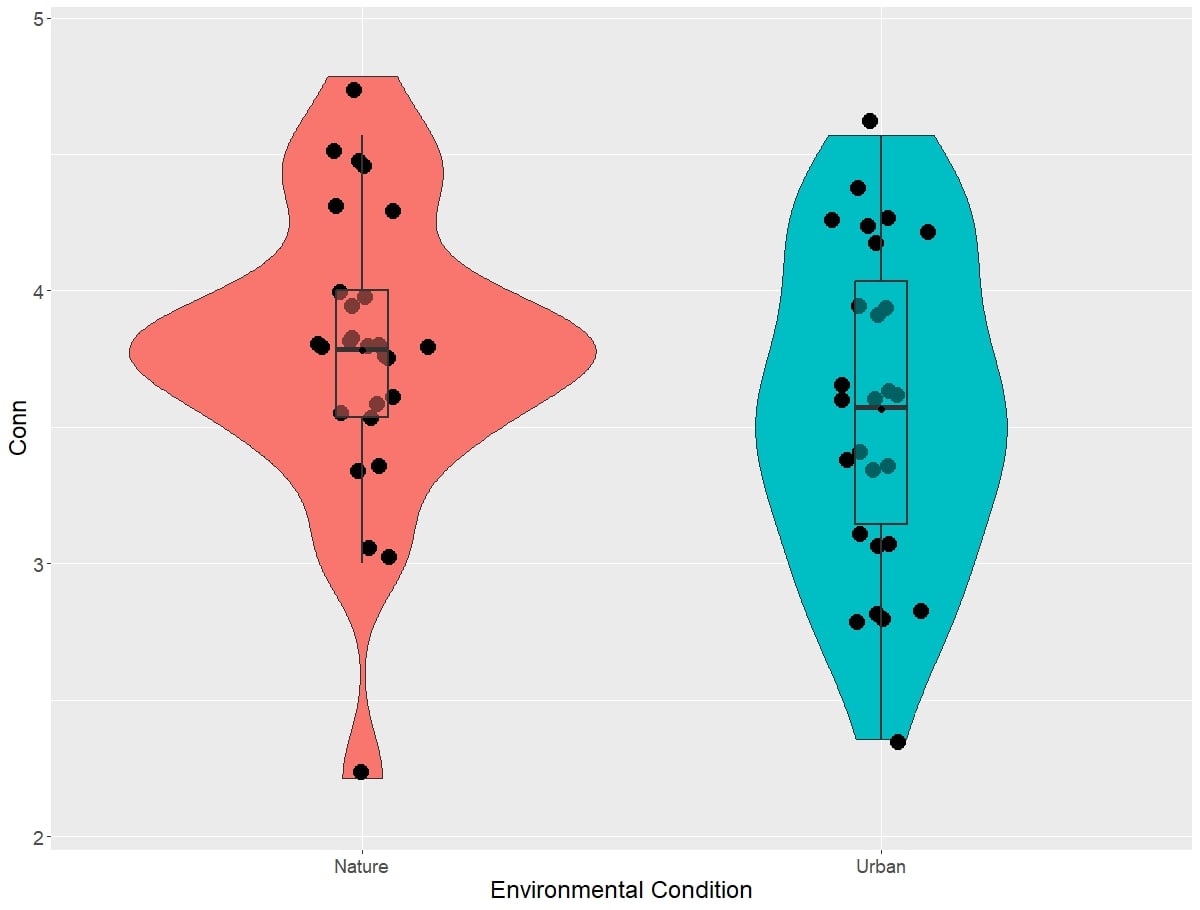
Figure 2: Self-reported connectedness to nature scores between environmental conditions
Classic Stroop Task Results
Reaction time results from the Classic Stroop task can be visualized in Figure 3, while difference scores on this task broken down by environment type are visualized in Figure 4. Linear mixed models revealed no significant main effect of environment type on difference scores on the Classic Stroop task (χ2(1)=1.27, p=0.260). Adding score on the Connectedness to nature scale as a covariate in the model did not significantly influence model results (χ2(1)=0.53, p=0.465). Next, the effect of session number on the difference scores was assessed. Linear mixed models did not reveal a significant main effect of session on the Classic Stroop task (χ2(1)=3.50, p=0.0623). Adding the score on the connectedness to nature scale as a covariate factor in the model did not influence the model results (χ2(1)=1.19, p=0.280). Linear mixed models did not find the interaction of session and environment type to have a significant effect on the difference scores on the Classic Stroop Task(χ2(1)=0.946, p=0.354). Again, adding the connectedness to nature scale as a covariate did not have a significant impact on the model results (χ2(1)=0.442, p=0.375).
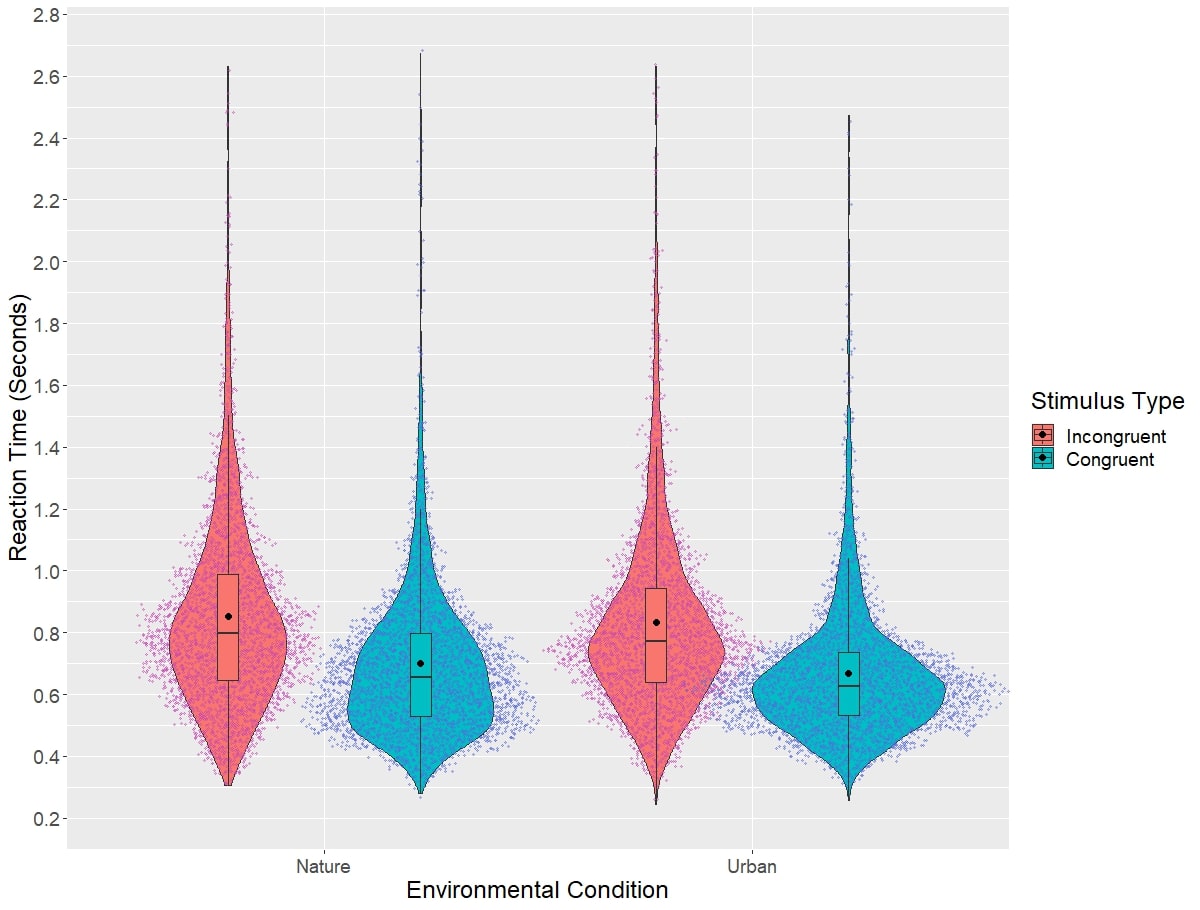
Figure 3: Results reaction times to the Classic Stroop task, broken down by stimulus type and testing environment.
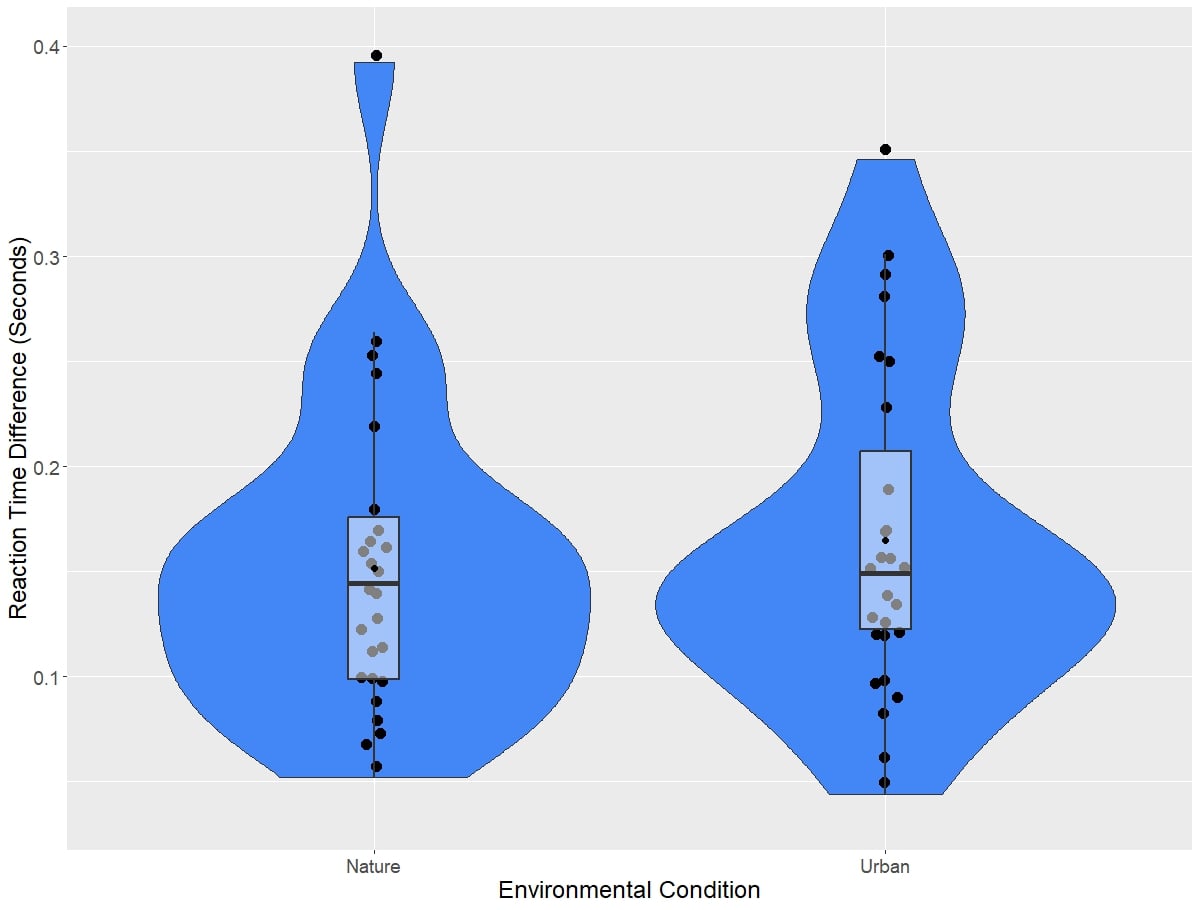
Figure 4: Difference scores on the Classic Stroop task, broken down by environmental condition.
Emotional Stroop Task Results
Reaction time results from the Emotional Stroop task can be visualized in Figure 5, while difference scores on this task broken down by environment type are visualized in Figure 6. Linear mixed models revealed no significant main effect of the environment type on the difference scores on the Emotional Stroop Task (χ2(1)=2.86, p=0.101). Additionally, linear mixed models did not reveal a significant main effect of session on the difference scores of the Emotional Stroop Task (χ2(1)=0.273, p=0.612). Linear mixed models also did not find a significant interaction between environment type and session on the Emotional Stroop Task difference scores (χ2(1)=3.91, p=0.390). Adding connectedness to nature scores did not significantly impact the models for environment (χ2(1)=3.71, p=0.159), session (χ2(1)=1.66, p=0.539), or the interaction of environment and session (χ2(1)=0.842, p=0.408).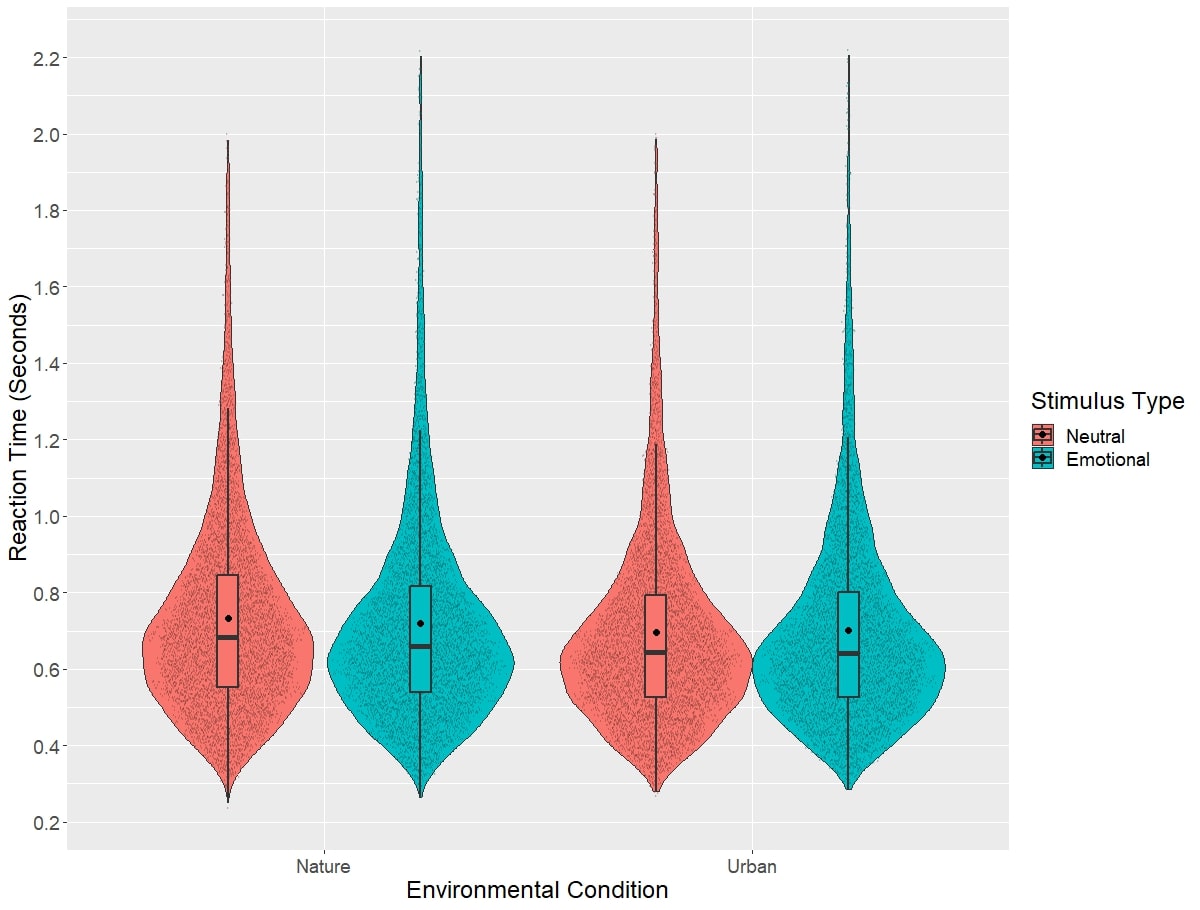
Figure 5: Reaction times to the Emotional Stroop task, by stimulus type and environment.
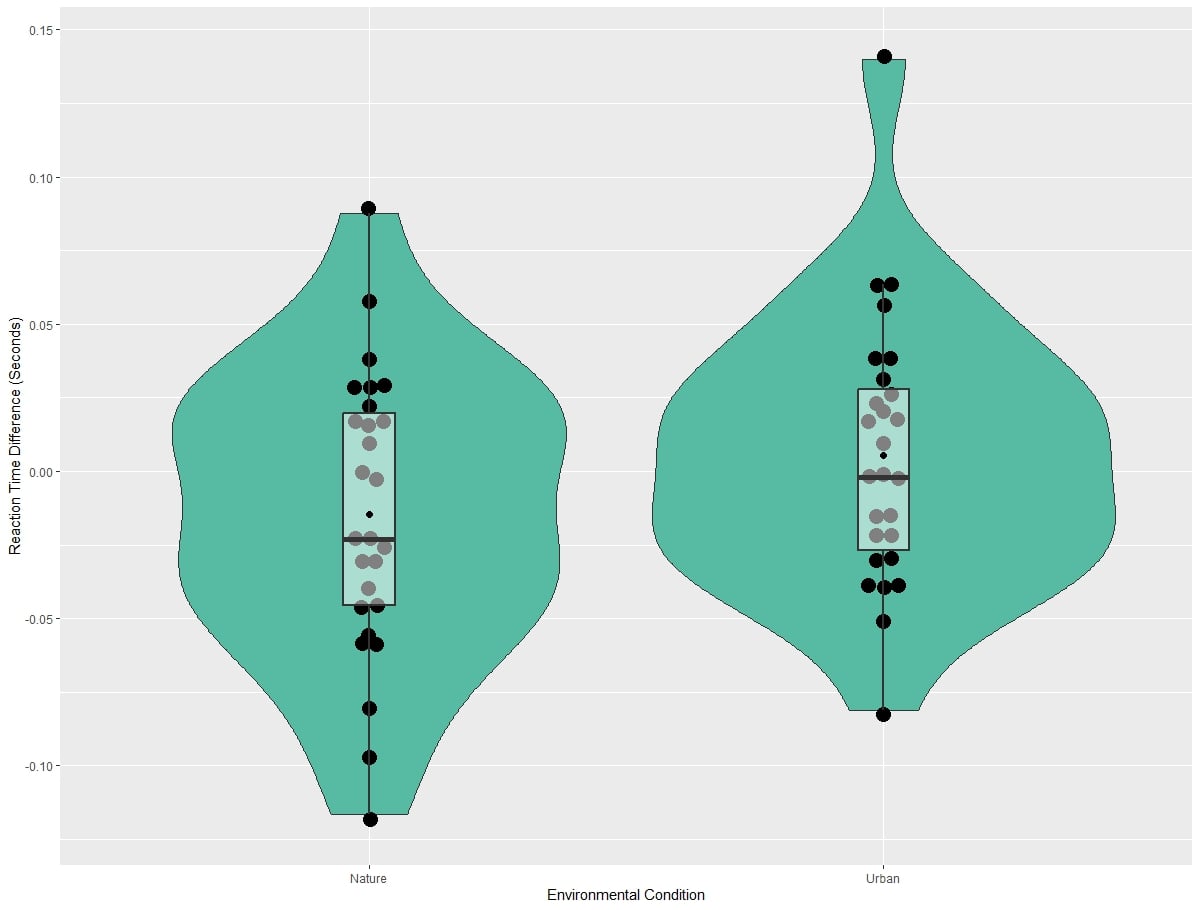
Figure 6: Difference scores for the Emotional Stroop task by environment.
Discussion
The present study sought to explore whether or not immersion in nature influenced cognitive control (as measured via performance on the Classic Stroop Task) and emotional inhibitory control (measured via performance on the Emotional Stroop Task), and how these relationships may have been influenced by each participants’ self-reported connectedness to nature. Both of these cognitive processes are thought to be facets of directed, executive attention and thus are thought to be improved by immersion in nature per Attention Restoration Theory (Kaplan, 1995).
Our analysis did not reveal a significant effect of environment type on Classic Stroop or Emotional Stroop task performance. Furthermore, neither the connectedness to nature (CNS) scores nor session number had a significant impact on the performance of these tasks. Our analysis however did find that CNS scores were significantly higher in the nature condition compared to the urban condition.
Our hypothesis that exposure to natural environments would result in significant benefits to the Stroop and Emotional Stroop task was not supported by the results. There are a number of factors why this may have been the case. First, although the locations that were selected in both conditions were intended to be free from distractions, elements such as nearby wildlife, sun and ambient sounds may have still distracted participants and affected overall results. Furthermore, since only the difference scores on the tasks were analyzed, it is possible that subjects made similar improvements in responding to both the congruent and incongruent trials in the classic Stroop task, and to the neutral and emotional trials in the Emotional Stroop task. Such improvements would not result in a change of difference scores, and thus would not be captured by the analyses performed. Finally, although score on the Connectedness to Nature scale has been previously found to partially explain benefits from exposure to nature (Mayer et al., 2008), these correlations are not universally observed and may be a proxy to related influences such as familiarity with natural environments which may reasonably impact the restorative benefits of nature. This is supported by the greater variability in reaction times in the natural condition which can be seen in Figure 3 and Figure 6. Finally, it is possible that the sample size of 27 participants resulted in our analyses being underpowered to detect more minute effects. We did observe a significant effect of environment type on Connectedness to Nature scores, which has important implications. Although the study sought to uncover benefits to cognitive and emotional processing in nature, higher levels of connectedness to nature are still a notable benefit. Increased CNS scores are correlated with higher engagement with natural environments, increased pro-environmental behaviors, and greater sense of stewardship for natural environments (Martin et al., 2020; Whitburn et al., 2019).
Conclusion
While this study did not find significant evidence that immersion in natural environments improves cognitive control in response inhibition or emotional response inhibition, benefits to self-reported connectedness to nature were found. Furthermore, the sample size of this study may have resulted in underpowered analyses that were not able to detect smaller effects which may be present. Future studies should seek to expand on the impact of immersion in nature on subjective feelings toward nature. Additionally, replicating these results with a larger sample size may implicate differences in restorative effects of nature based on the specific natural environment subjects are exposed to.
References
Atchley R. A., Strayer D.L., & Atchley, P. (2012). Creativity in the Wild: Improving Creative Reasoning through Immersion in Natural Settings. PLoS ONE, 7(12): e51474. https://doi.org/10.1371/journal.pone.0051474
Bailey, A. W., Allen, G., Herndon, J., & Demastus, C. (2018). Cognitive Benefits of walking in natural versus built environments. World Leisure Journal, 60(4), 293-305. https://doi.org/10.1080/16078055.2018.1445025
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software, 67, 1–48. https://doi.org/10.18637/jss.v067.i01
Ben-Haim M.S., Williams P., Howard, Z., Mama, Y., Eidels, A., & Algom, D. (2016). The Emotional Stroop Task: Assessing Cognitive Performance under Exposure to Emotional Content. Journal of Visualized Experiments, 29(112). doi: 10.3791/53720.
Berman, M. G., Kross, E., Krpan, K. M., Askren, M. K., Burson, A., Deldin, P. J., … & Jonides, J. (2012). Interacting with nature improves cognition and affect for individuals with depression. Journal of affective disorders, 140(3), 300-305.
Bratman, G. N., Hamilton, J. P., Hahn, K. S., Daily, G. C., & Gross, J. J. (2015). Nature experience reduces rumination and subgenual prefrontal cortex activation. Proceedings of the National Academy of Sciences, 112(28), 8567–8572. https://doi.org/10.1073/pnas.1510459112
Crossfield, E., Damian, M.F. (2021) The role of valence in word processing: Evidence from lexical decision and emotional Stroop tasks. Acta Psychologica, 218(2021). https://doi.org/10.1016/j.actpsy.2021.103359
Golembiewski, J. (2019). Neuroscience of mental illness in the city. In Bhugra, D., Ventriglio, A., McCay, L., Castaldelli-Maia, J.(Eds.), Urban Mental Health (108-120). Oxford University Press. https://doi.org/10.1093/med/9780198804949.003.0009
Hartig, T., Mang, M., & Evans, G.W. (1991). Restorative effects of Natural Environment Experiences. Environment and Behavior, 23(1), 3-26. https://doi- org.ezproxy.lib.utah.edu/10.1177/0013916591231001
Hasshim, N., & Parris, B. A. (2021). The role of contingency and correlation in the Stroop task.
Quarterly Journal of Experimental Psychology, 74(10), 1657–1668. https://doi.org/10.1177/17470218211032548
Johnson, D.R., (2009). Goal-directed attentional deployment to emotional faces and individual differences in emotional regulation. Journal of Research in Personality, 43(1), 8-13. https://doi.org/10.1016/j.jrp.2008.09.006.
Kaplan, S., and Berman, M.G., (2010). Directed attention as a common resource for executive functioning and self-regulation. Perspectives on Psychological Science, 5,,43-57. https://doi.org/10.1177/1745691609356784’
Kaplan, S. (1995). The restorative benefits of nature: Toward an integrative framework. Journal of Environmental Psychology, 15(3), 169–182
Koselka, E.P.D., Weidner, L.C., Minasov, A., Berman, M.G., Leonard, W.R., Santoso, M.V., de Brito, J.N., Pope, Z.C., Pereira, M.A., & Horton, T.H. (2019). Walking Green: Developing an Evidence Base for Nature Prescriptions. International Journal of Environmental Research and Public Health 2019, 16(22). https://doi.org/10.3390/ijerph16224338
Lambert, K. G., Nelson, R. J., Jovanovic, T., & Cerdá, M. (2015). Brains in the city: Neurobiological effects of urbanization. Neuroscience & Biobehavioral Reviews, 58, 107-122. https://doi.org/10.1016/j.neubiorev.2015.04.007
Linnell, K.J., S. Caparos, J.W. de Fockert, and J. Davidoff. (2013). Urbanization decreases attentional engagement. Journal of Experimental Psychology: Human Perception and Performance 39(5): 1232–1247. https://doi.org/10.1037/a0031139
LoTemplio, S. B., Scott, E. E., McDonnell, A. S., Hopman, R. J., Castro, S. C., McNay, G. D., McKinney, T. L., Greenberg, K., Payne, B. R., & Strayer, D. L. (2020). Nature as a potential modulator of the error-related negativity: a registered report. International Journal of Psychophysiology, 156, 49–59. https://doi.org/10.1016/j.ijpsycho.2020.06.014
Martin, L., White, M., Hunt, A., Richardson, M., Pahl, S., & Burt, J. (2020). Nature contact, nature connectedness, and associations with health, wellbeing and pro-environmental behaviors. Journal of Environmental Psychology, 68. https://doi.org/10.1016/j.jenvp.2020.101389
Mayer, F. S., & Frantz, C. M.(2004). The connectedness to nature scale: A measure of individuals’ feeling in community with nature. Journal of Environmental Psychology, 24(4), 503-515.
Mayer, F. S., Frantz, C. M., Bruehlman-Senecal, E., & Dolliver, K. (2009). Why Is Nature Beneficial?: The Role of Connectedness to Nature. Environment and Behavior, 41(5), 607–643. https://doi.org/10.1177/0013916508319745
Scott, E. E., McDonnell, A. S., LoTemplio, S. B., Uchino, B. N., & Strayer, D. L. (2021). Toward a unified model of stress recovery and cognitive restoration in nature. Parks Stewardship Forum, 37(1). https://doi.org/10.5070/p537151710
Straub, E.R., Schmidts, C., Kunde, W., Zhang, J., Kiesel, D., & Diganth, D. (2021). Limitation of cognitive control on emotions distraction – Congruency in the Color Stroop task does not modulate the Emotional Stroop Task. Cognitive, Affective, & Behavioral Neuroscience, 22, 21-41.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18, 643–662.
The World Bank. (2023, April 03). “Urban Development”. The World Bank. https://www.worldbank.org/en/topic/urbandevelopment/overview#:~:text=Today%2C%20some%2056%25%20of%20the,people%20will%20live%20in%20cities
Whitburn, J., Linklater, W., & Abrahamse, W. (2019). Meta-analysis of human connection to nature and proenviornmental behavior. Conservation Biology, 34(1), 180-191. https://doi.org/10.1111/cobi.13381
