Retinal Wiring
Jim Hutchins
Objective 5: Describe how each of the five neuronal types of the retina contribute to visual processing.
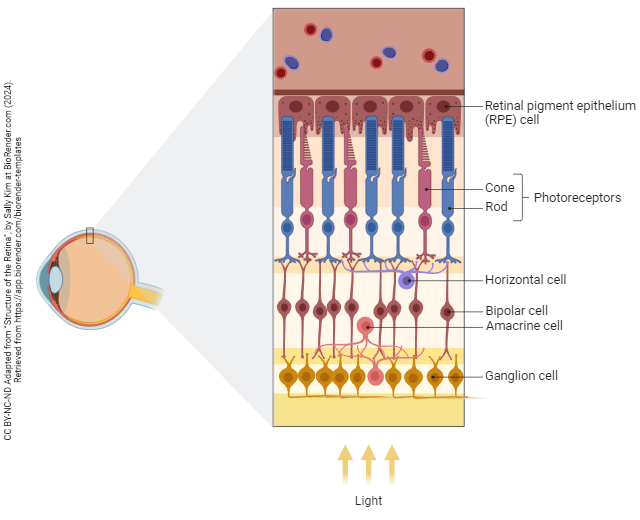 There are five types of neurons in the retina:
There are five types of neurons in the retina:
- photoreceptors, the neurons which transduce photons into electrical potentials and release neurotransmitter onto the processing cells;
- horizontal cells, neurons that share information between neighboring photoreceptors;
- bipolar cells, which convey information between photoreceptors and ganglion cells;
- amacrine cells, neurons that share information between neighboring bipolar cells;
- ganglion cells, the output cells of the retina.
There is also a glial cell, which resembles an astrocyte, called a Müller cell.
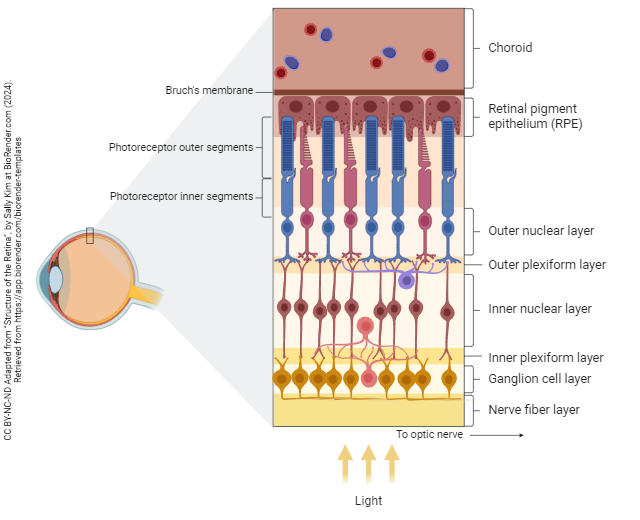
The blood supply of the outer retina (i.e. photoreceptors) comes from the choroid, also called the choriocapillaris. The choroid is part of a layer called the vascular tunic or uvea and is continuous with the ciliary body and iris.
Layers of the Neural Retina
The retinal pigment epithelium (RPE) passes nutrients from the choriocapillaris to the photoreceptor outer segments. Transduction in photoreceptors produces a huge energy demand which can only be met if there is a tight connection between the RPE and photoreceptors. In a detached retina, this tight connection is lost and the neural retina dies. The nuclei of photoreceptors are found in the outer nuclear layer (ONL). Their synaptic terminals pass information to horizontal cells and bipolar cell dendrites in the outer plexiform layer (OPL, also called the outer synaptic layer). Bipolar cell bodies and some amacrine cell bodies are found in the inner nuclear layer (INL). The slightly mis-named ganglion cell layer (GCL) contains ganglion cell bodies and some amacrine cell bodies. The axons of ganglion cells form the nerve fiber layer (NFL).
“Straight-Through” Information Processing by Retinal Cells
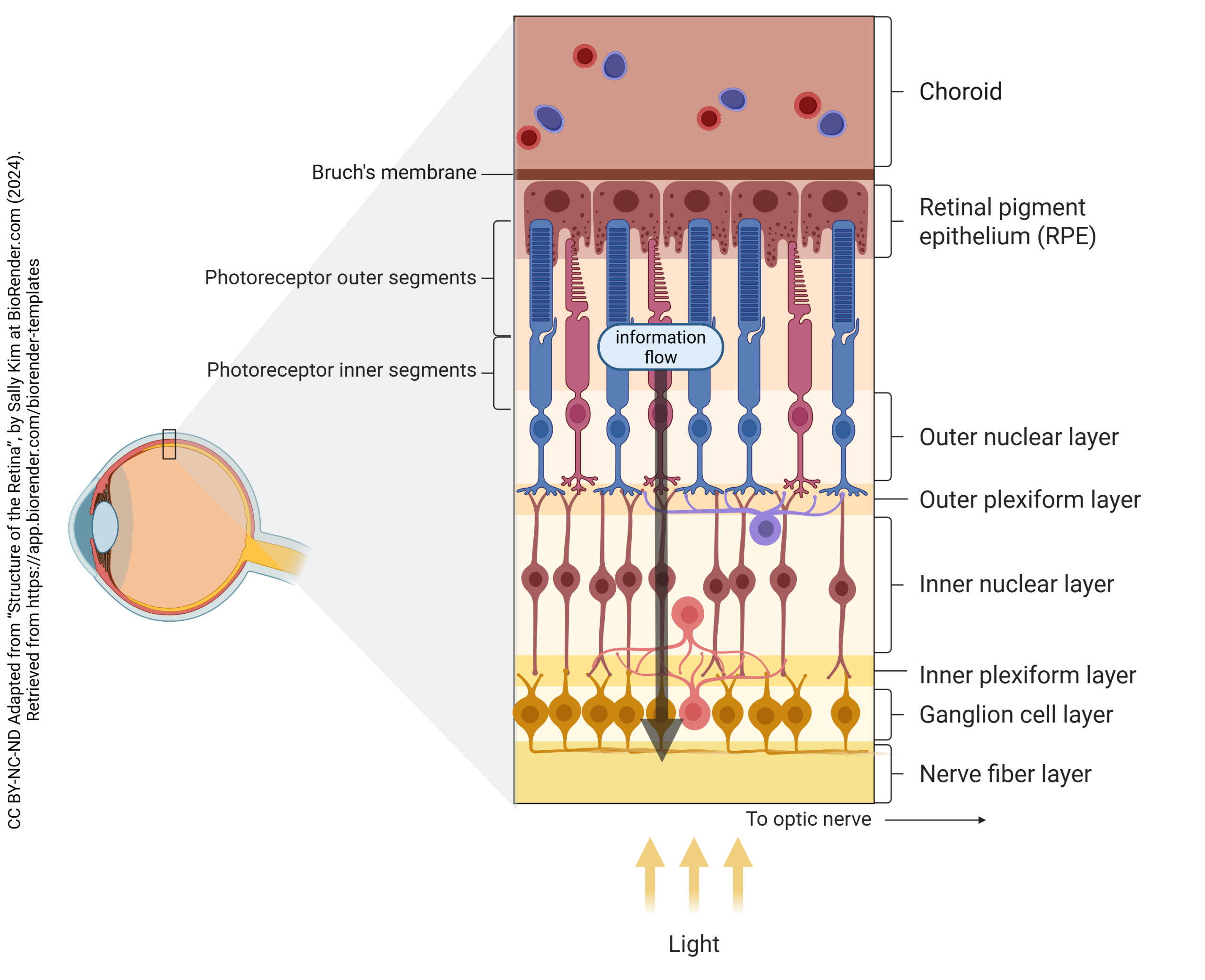
Photoreceptors, bipolar cells, and ganglion cells can form an information processing chain that is called the straight-through pathway.
Lateral Information Processing by Retinal Cells
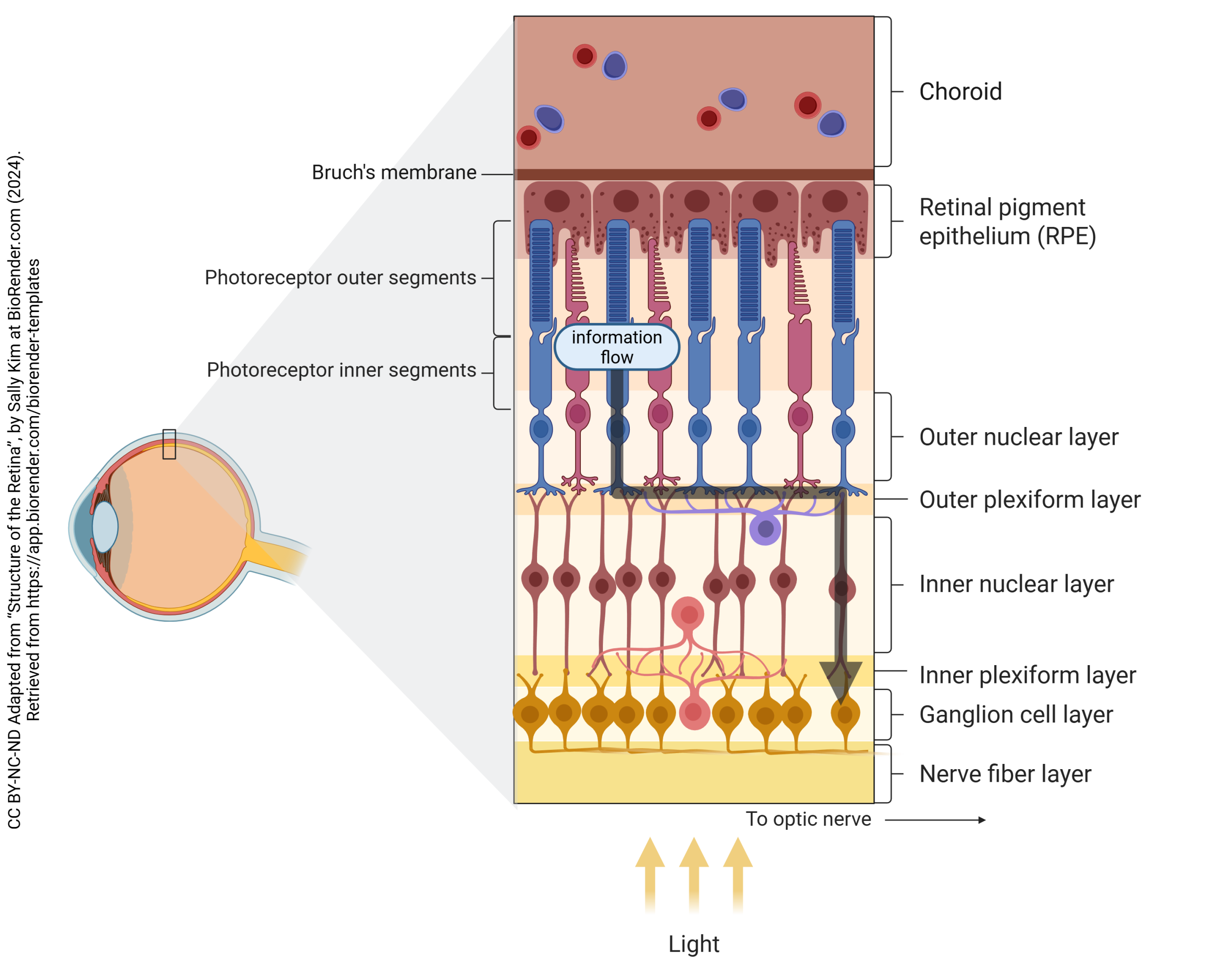
The retina can process information “sideways”, sharing information with neighboring cells through the horizontal cell and amacrine cell network. This is called the lateral pathway.
As the name suggests, horizontal cells pass information horizontally in the retina, telling neighboring photoreceptors what is going on nearby. They have few discernable synapses, and both receptors and neurotransmitter release mechanisms are not very visible but must exist in the OPL.
Bipolar cells gather together information coming from photoreceptors and horizontal cells. While all photoreceptors release glutamate, the receptors on bipolar cells fall into two categories.
Receptive Fields of Bipolar Cells
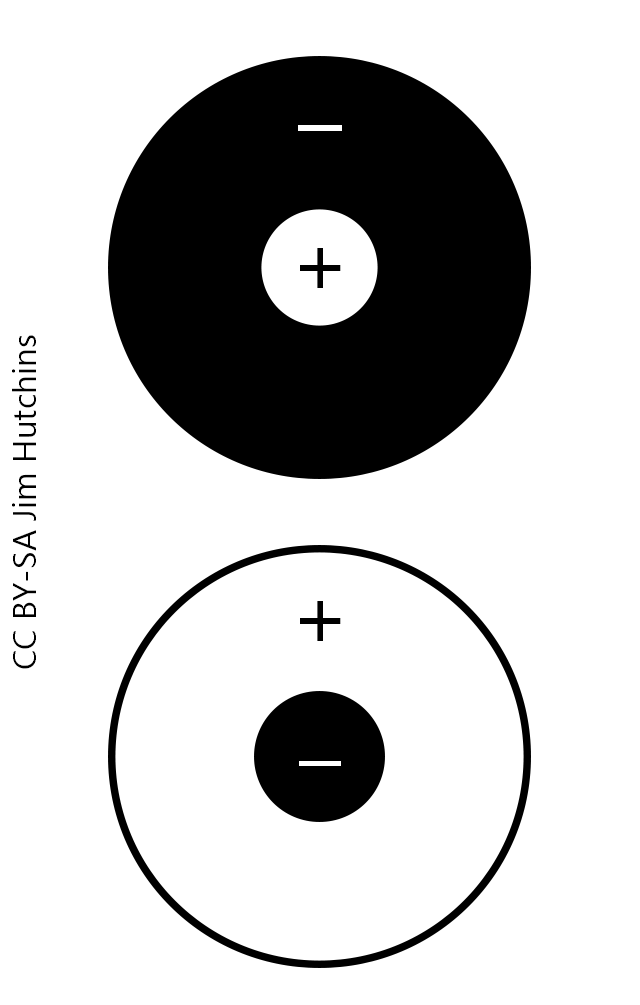
The receptive field of a bipolar cell is the complete set of points in visual space that cause activation of a particular bipolar cell. A bipolar cell which is close to the fovea has no knowledge of what is going on in the peripheral retina near the ciliary body, and vice versa. Lateral interactions are hyperlocal and used only to set up the individual bipolar’s receptive field. The receptive field of a bipolar cell is an exact overlay, then, of the bipolar cell’s dendritic tree.
Bipolar cells have center-surround receptive fields. This means that in the center of the tree, nearest the “trunk”, one response predominates while in the faraway branches of the tree the opposite type of synapse is found.
Center-surround receptive fields predominate in the retina, in the lateral geniculate nucleus, and in the initial processing steps in visual cortex.
Sign-Conserving vs Sign-Inverting Synapses in the Retina
Bipolar cells are called “ON-bipolars” if they are excited by light applied to the photoreceptor which lies directly above them. Bipolar cells are called “OFF-bipolars” if they are hyperpolarized by light applied to the photoreceptor which lies directly above them.
In the center of an ON-bipolar receptive field (top circles in the diagram), there are glutamate receptors of the metabotropic mGluR6 type. When mGluR6 receptors are occupied by glutamate, they cause a hyperpolarization of the bipolar cell. Conversely, when the mGluR6 receptors are unoccupied, they allow the bipolar cell membrane potential to rise to threshold for neurotransmitter release from the synaptic terminals. Light causes a decrease in glutamate release and fewer mGluR6 receptors are occupied. This is why the top diagram shows a plus sign in the center circle (depolarization of the bipolar cell). Because the hyperpolarization of photoreceptors causes a depolarization of the bipolar cell on the other side of the synapse, this is called a “sign-inverting” synapse.
Sign-inverting synapses would be called “inhibitory” in a different context.
Some bipolar cells are classified as “OFF-bipolars” because in the center of their receptive fields, they respond to light with a hyperpolarization (bottom center circle). This is symbolized by a minus sign. These are “sign-conserving” synapses. A hyperpolarization in the photoreceptor (i.e. in the light) results in a hyperpolarization in the center of the bipolar cell’s receptive field. This is mediated by NMDA and non-NMDA type glutamate receptors. When glutamate is plentiful, as in the dark, they result in a depolarization of bipolar cells with postsynaptic elements directly underneath the photoreceptor. When glutamate is scarce, as in the light, they result in a hyperpolarization of bipolar cells with postsynaptic elements directly underneath the photoreceptor.
Sign-conserving synapses would be called “excitatory” in a different context.
ON-bipolar cells (top circles in the diagram) have sign-inverting synapses on their central dendrites, and sign-conserving synapses on the faraway parts of the dendritic tree. This gives them a center-surround receptive field organization.
OFF-bipolar cells (bottom circles in the diagram) have sign-conserving synapses on their central dendrites, and sign-inverting synapses on the faraway parts of the dendritic tree. They also have a center-surround receptive field organization.
How Receptive Fields Are Built
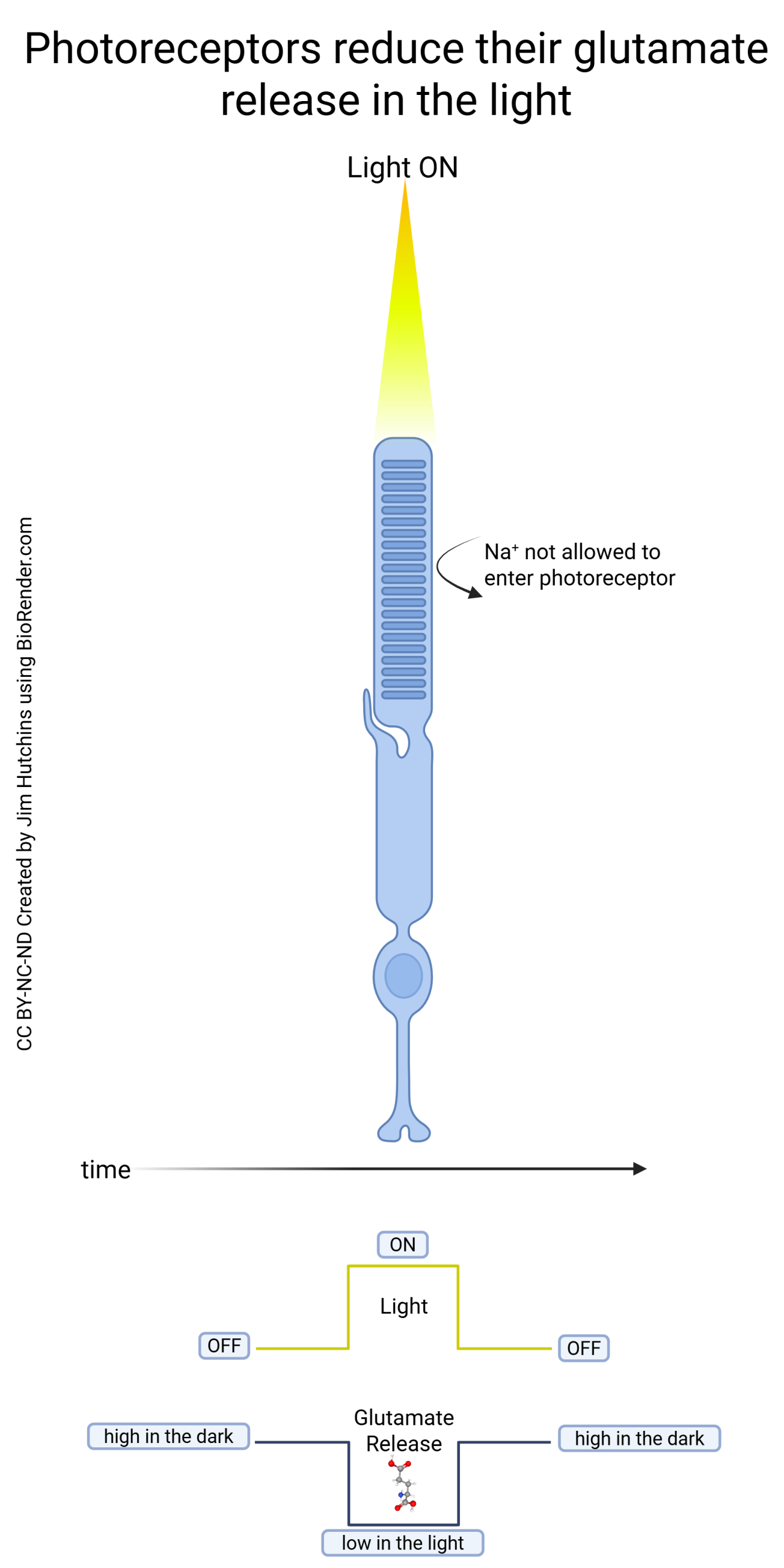
Remember from the “Visual Transduction” chapter that when light shines on an individual photoreceptor, that photoreceptor hyperpolarizes and releases less glutamate onto horizontal and bipolar cells.
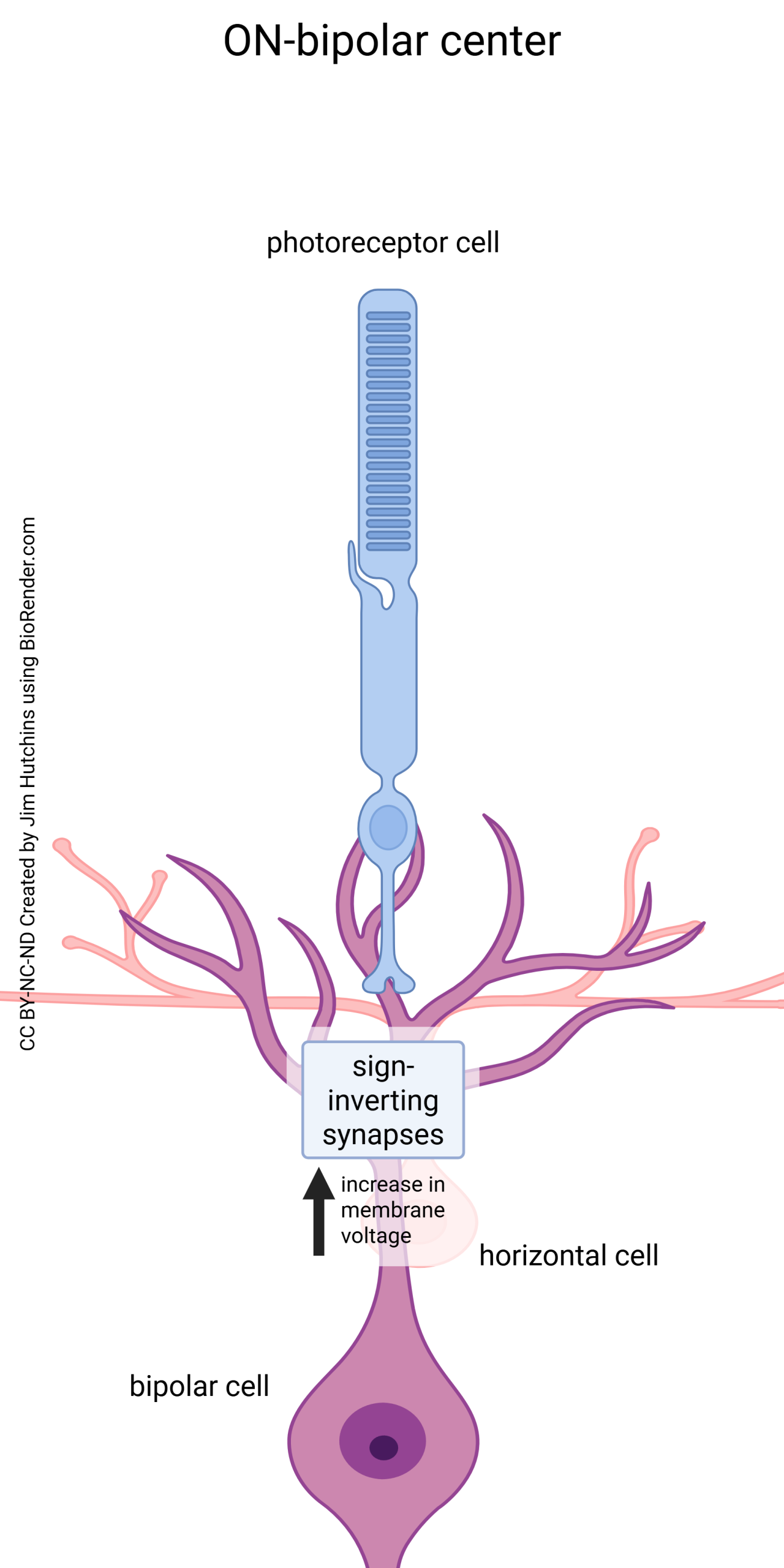
In the center of the receptive field, we find sign-inverting synapses that convert this hyperpolarization in the photoreceptor to a depolarization in the central dendrites of a bipolar cell.
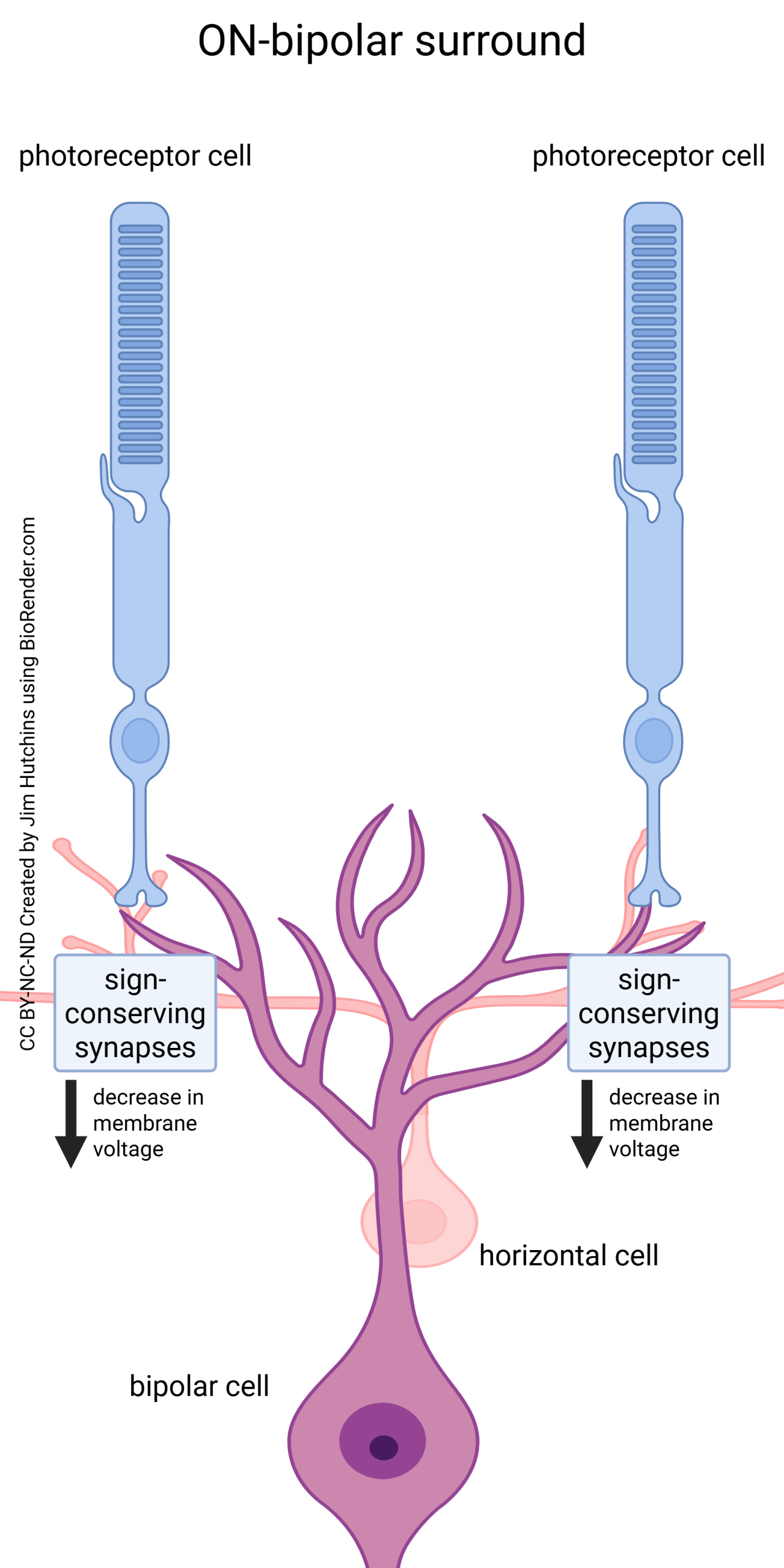
In the surround of the receptive field, we find sign-conserving synapses, where a hyperpolarization in the photoreceptor results in a hyperpolarization in the outlying dendrites of the bipolar cell.
Media Attributions
- Structure of the retina cell types © Sally Kim adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Structure of the retina layers © Sally Kim adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Straight-through processing in the retina © Sally Kim adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Lateral processing in the retina © Sally Kim adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Center surround receptive fields © Jim Hutchins is licensed under a CC BY-SA (Attribution ShareAlike) license
- How photoreceptors respond to light © Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Constructing an ON bipolar cell center © Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Constructing an ON bipolar cell surround © Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
