The Cross-Bridge Cycle and Excitation-Contraction Coupling
Caleb Bevan
Objective 4: Illustrate the cross-bridge cycle and excitation-contraction coupling in skeletal muscle.
Management of Electrical Activity in Skeletal Muscle
The action potential in a muscle initiates contraction through a series of chemical reactions and protein interactions.
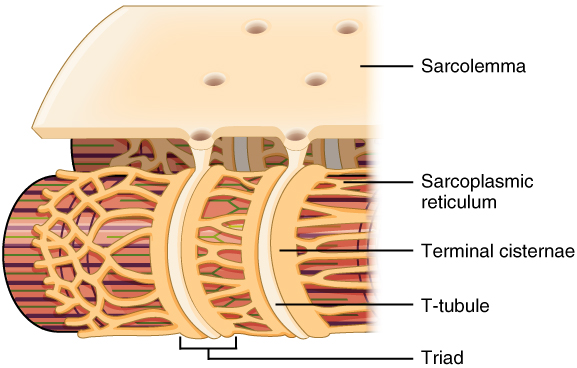
The action potential is initiated at the neuromuscular junction, as described in the last chapter. It then spreads over the surface of the muscle cell as described earlier for action potential propagation in a nerve axon.
Unlike the axon, however, the function of the muscle cell action potential is not to activate a synapse, but rather to initiate contraction over the entire muscle fiber all at once.
To do this, structures called transverse tubules (T-tubules) dive deep inside the muscle fiber. As the name implies, they are arranged perpendicular to the surface of the sarcolemma. Deep inside the muscle fiber, they form a three-part arrangement called a triad with two saddlebags of sarcoplasmic reticulum.

Remember that sarcoplasmic reticulum is a special kind of smooth endoplasmic reticulum. In this case, the function of the sarcoplasmic reticulum is to act as a storage system for calcium ions (Ca2+).
Calcium Release from Sarcoplasmic Reticulum
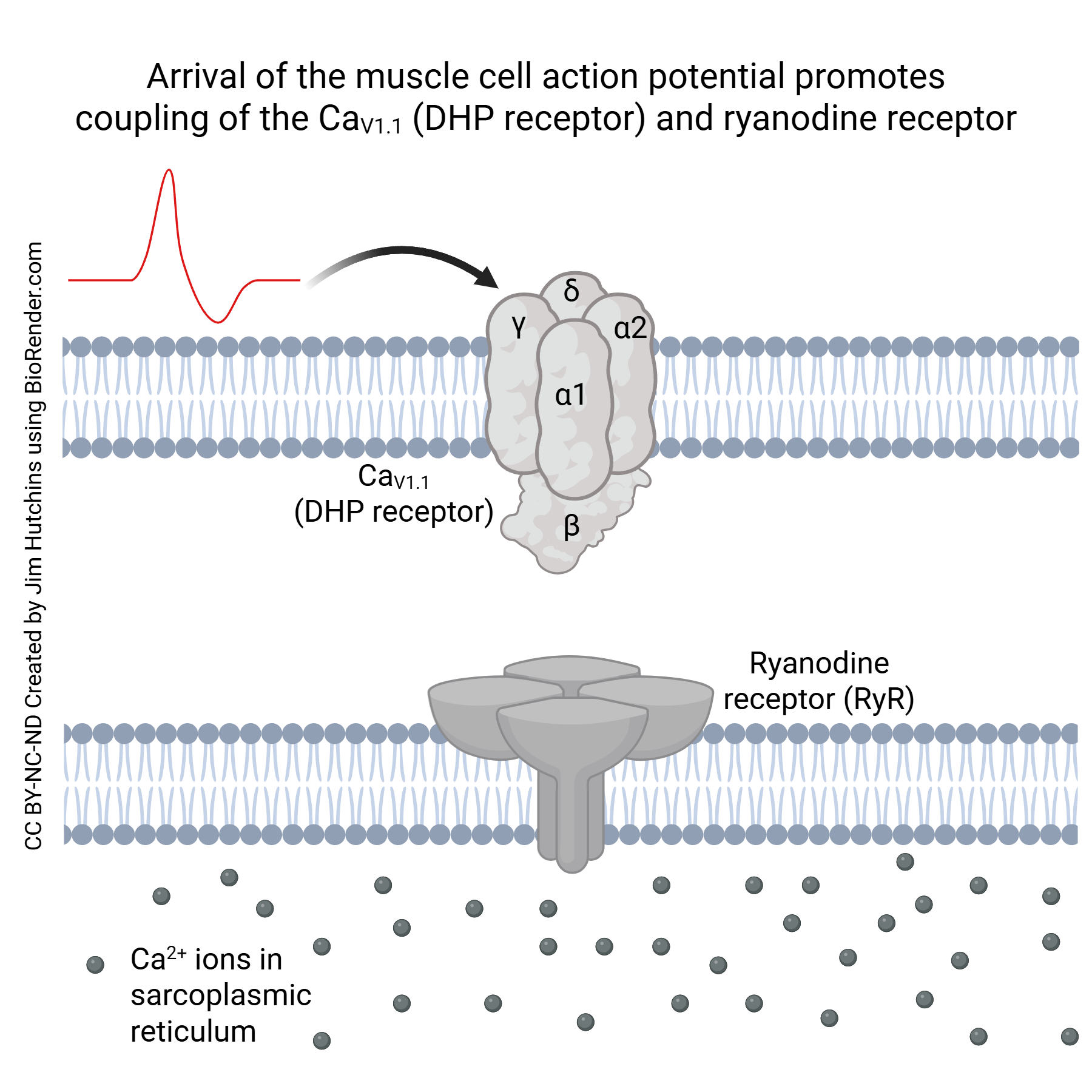 At the triad, the voltage-gated calcium channel CaV1.1 (also called the dihydropyridine receptor or DHP receptor) is found on the sarcolemma (i.e. cell membrane of the muscle cell). The ryanodine receptor (RyR) is embedded in the membrane sarcoplasmic reticulum. When the muscle cell is at its resting potential, these receptors are near each other but uncoupled.
At the triad, the voltage-gated calcium channel CaV1.1 (also called the dihydropyridine receptor or DHP receptor) is found on the sarcolemma (i.e. cell membrane of the muscle cell). The ryanodine receptor (RyR) is embedded in the membrane sarcoplasmic reticulum. When the muscle cell is at its resting potential, these receptors are near each other but uncoupled.
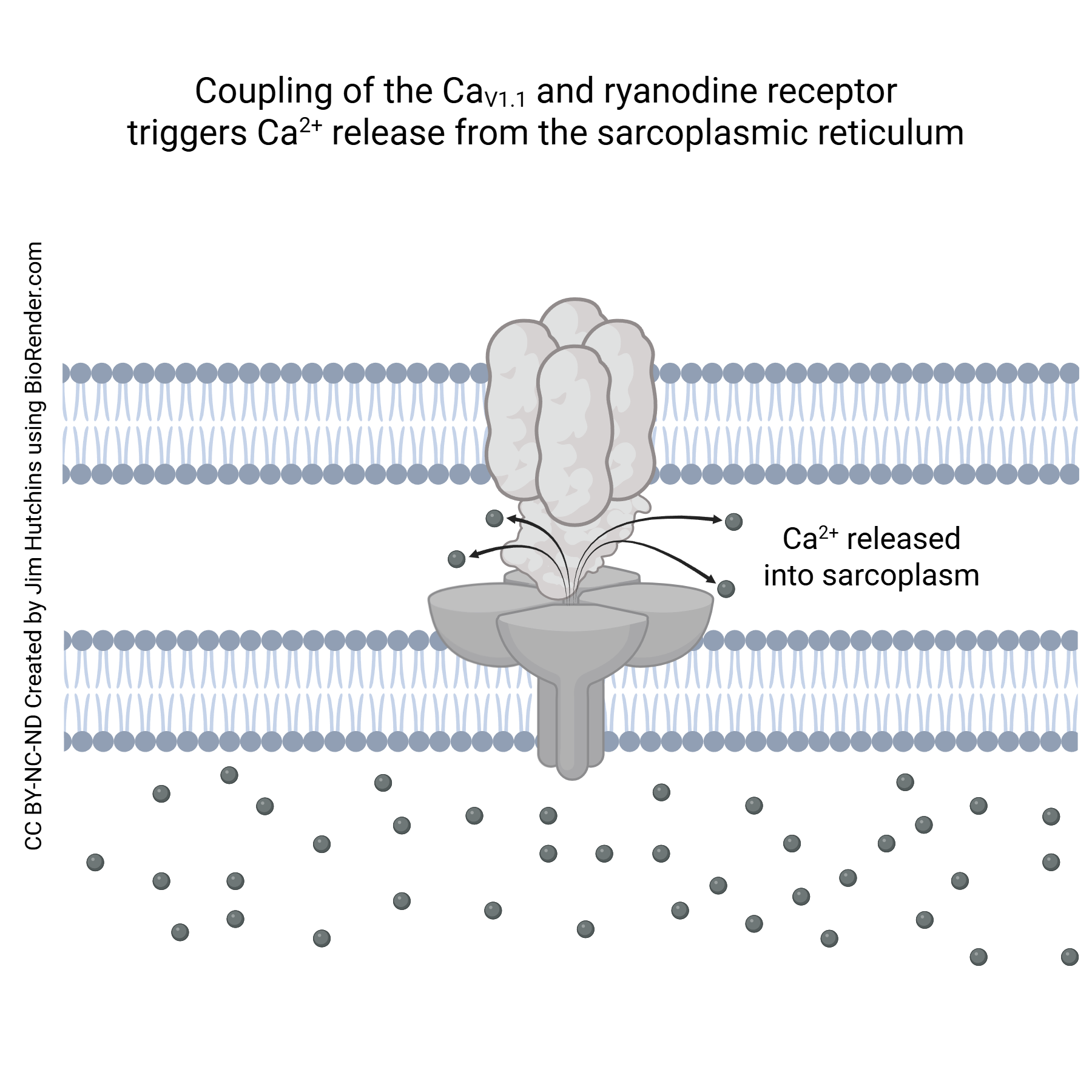 A muscle cell action potential transduce the voltage change into Ca2+ release. The DHP receptor is an L-type voltage-gated Ca2+ channel. The RyR is linked to this voltage-gated Ca2+ channel; when the DHP receptor changes shape in response to the voltage change, it causes a shape change in the RyR and this configuration of the RyR allows Ca2+ to leave the sarcoplasmic reticulum.
A muscle cell action potential transduce the voltage change into Ca2+ release. The DHP receptor is an L-type voltage-gated Ca2+ channel. The RyR is linked to this voltage-gated Ca2+ channel; when the DHP receptor changes shape in response to the voltage change, it causes a shape change in the RyR and this configuration of the RyR allows Ca2+ to leave the sarcoplasmic reticulum.
The Ca2+ released from the sarcoplasmic reticulum interacts with troponin, a protein found only in muscle cells. When Ca2+ binds to troponin, it changes its shape, and pushes aside tropomyosin. Normally, tropomyosin covers the myosin binding sites on the actin molecule, preventing myosin heads from attaching and pulling in on the actin. When Ca2+ is present, and troponin shoves on tropomyosin, the myosin-binding site on the actin molecule is exposed, allowing the cross-bridge cycle to take place.
The Cross-Bridge Cycle
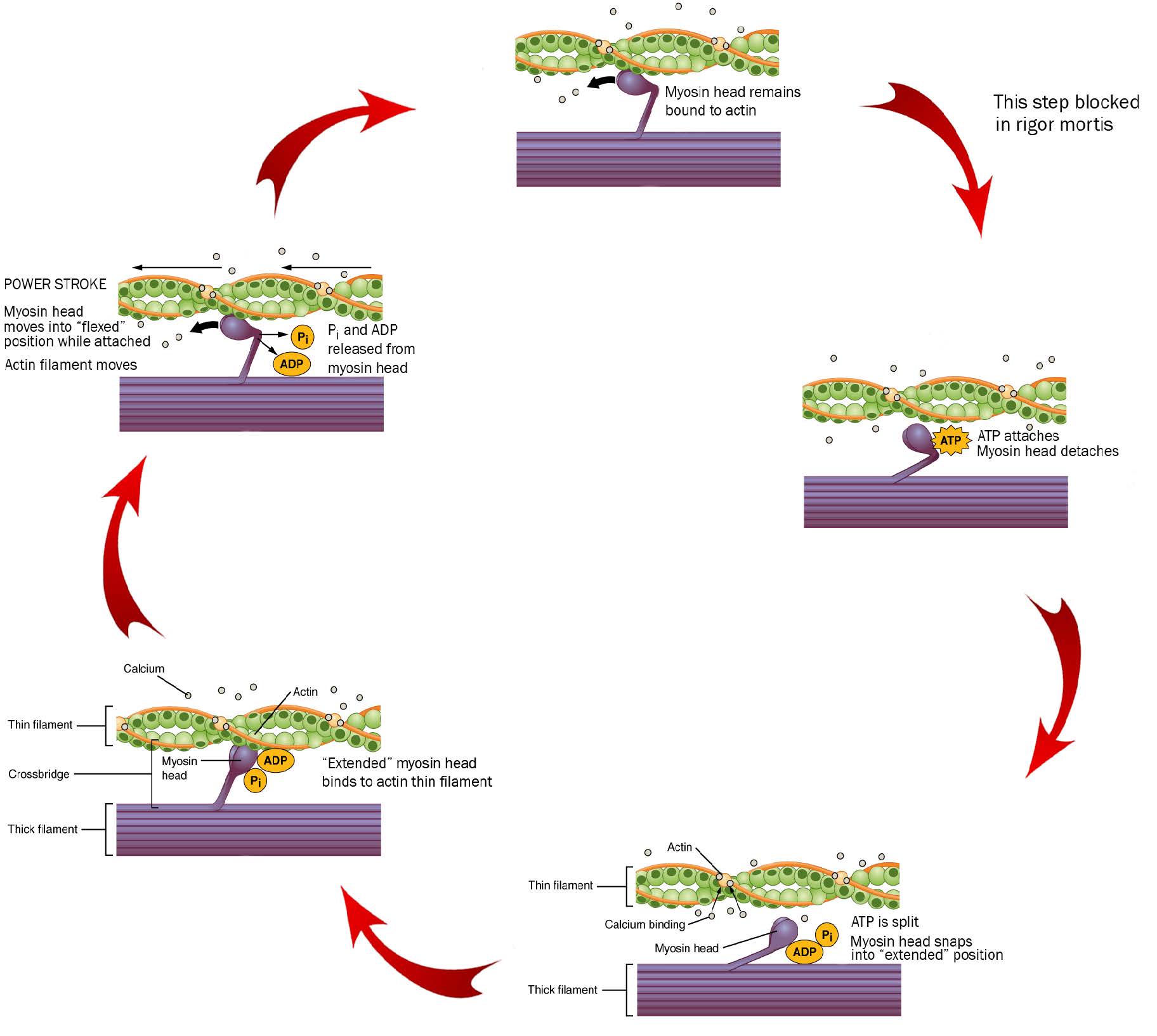 The cross-bridge cycle is (after all) a cycle, so we can start anywhere, but let’s start at the top of these diagrams, at 12 o’clock on an imaginary clock face.
The cross-bridge cycle is (after all) a cycle, so we can start anywhere, but let’s start at the top of these diagrams, at 12 o’clock on an imaginary clock face.
We start as the previous cross-bridge cycle has just completed, with the myosin head bound to the actin filament.
ATP is required for this process. When ATP binds to the myosin head, it releases the actin filament. People who are recently dead have no available ATP in their muscles. Because of this, we remain stuck at 12 o’clock in this diagram, with myosin bound irreversibly to actin. This biochemical condition is called rigor mortis when we see it in an entire human body.
At 3 o’clock, ATP binds to the myosin head and it releases the actin filament.
At 5 o’clock, ATP is split. The energy is stored in the myosin head for later use. Right now, the splitting of adenosine triphosphate (ATP) into adenosine diphosphate (ADP) and inorganic phosphate (Pi) results in a conformational change in the myosin head, converting it from a “flexed” (bent) position to an “extended” (straight) position.
At 8 o’clock, calcium comes in and causes a conformational change in troponin, which shoves the tropomyosin, exposing the myosin-binding site on actin molecules in the thin filament. Myosin, still in its “extended” position, grabs onto the actin thin filament.
At 10 o’clock, ADP and Pi leave the myosin head. Now we use the previously-stored energy from ATP. In the power stroke, the myosin head (still bound to actin) changes shape as ADP and Pi leave. As myosin heads “flex” (bend), the thick filament myosin heads slide the thin filament actin molecules along, generating force as the sarcomere shortens.
Notice the actin filament moving to the left in this diagram as the purple thick filament stays in one place. This is the connection to the sliding filament model seen earlier: actin and myosin are interdigitated in such a way that they can easily interact.
If the signal from the lower motor neuron stops, the calcium is actively pumped back into the sarcoplasmic reticulum. When this happens, troponin and tropomyosin return to their original location blocking actin binding sites. Without anything to bind to, myosin pulls away from the actin and the proteins mentioned earlier allow for the sarcomere to return to its normal, resting length.
Excitation-Contraction Coupling
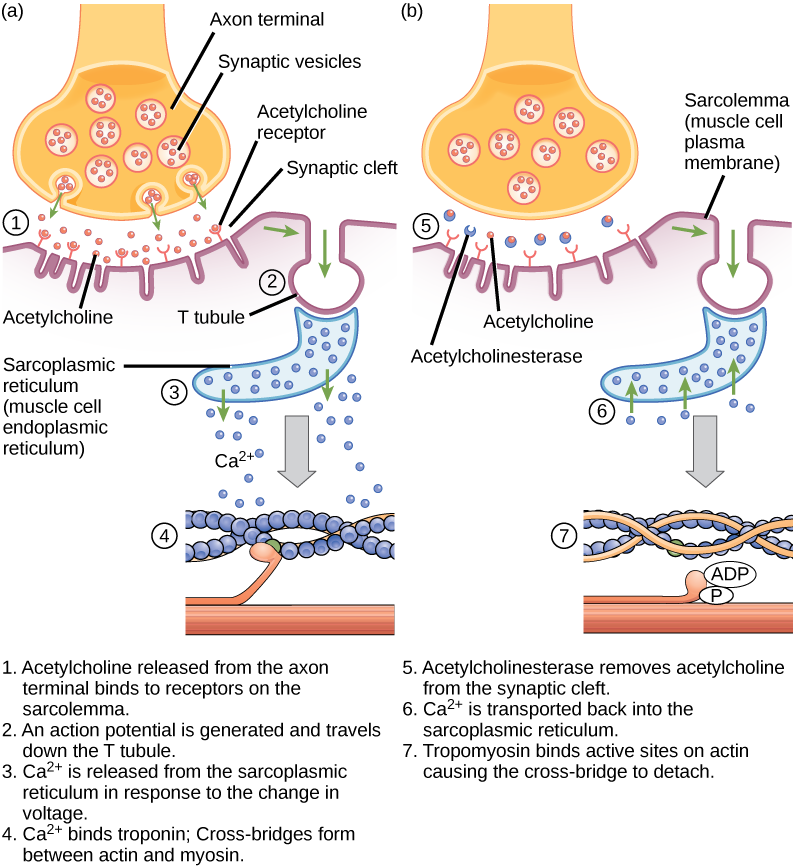 Now we are ready to put together all the steps involved in muscle cell contraction. Because this series of steps link excitation (the generation of a muscle cell action potential at the neuromuscular junction) to muscle cell contraction, they are called excitation-contraction coupling.
Now we are ready to put together all the steps involved in muscle cell contraction. Because this series of steps link excitation (the generation of a muscle cell action potential at the neuromuscular junction) to muscle cell contraction, they are called excitation-contraction coupling.
Media Attributions
- T-tubule © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license
- Toadfish muscle © Don W. Fawcett is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Calcium Release from Skeletal Muscle Sarcoplasmic Reticulum © Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Calcium Release from Skeletal Muscle Sarcoplasmic Reticulum © Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Cross-bridge cycle © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- Muscle depolarization © LibreTexts is licensed under a CC BY-SA (Attribution ShareAlike) license

{kind=link}