Neuronal Shape and Neuronal Information Processing
Jim Hutchins
Objective 3: Explain how the shape of neurons relates to their information processing properties.
The Shapes of Neurons
All of the neurons we have seen so far are multipolar. They have multiple processes (“sticky-outy-things”) coming out of the cell body.
Along with the multipolar neurons we’ve been looking at, there are two more shapes of neuron that deserve special mention.
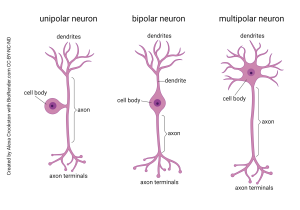
Bipolar neurons have two main processes, one emerging from each side of the cell body. As the name implies, the dendritic tree and the axon branches are close to mirror images of each other. They are found in places where information is relayed without much modification.
Unipolar neurons (also called pseudo-unipolar neurons) move the cell body off to the side, so the information is not processed or transformed as it bypasses the cell body. This type of neuron picks up information from the sensory surfaces of the skin and sends it, unmodified, to be dropped off in the spinal cord or brainstem. They are called unipolar because a single process emerges from the cell body.
In summary, to recognize the difference between multipolar, bipolar, and unipolar neurons, you should ask yourself: how many sticky-outy-things come out of the cell body?
Receiving and Processing Information
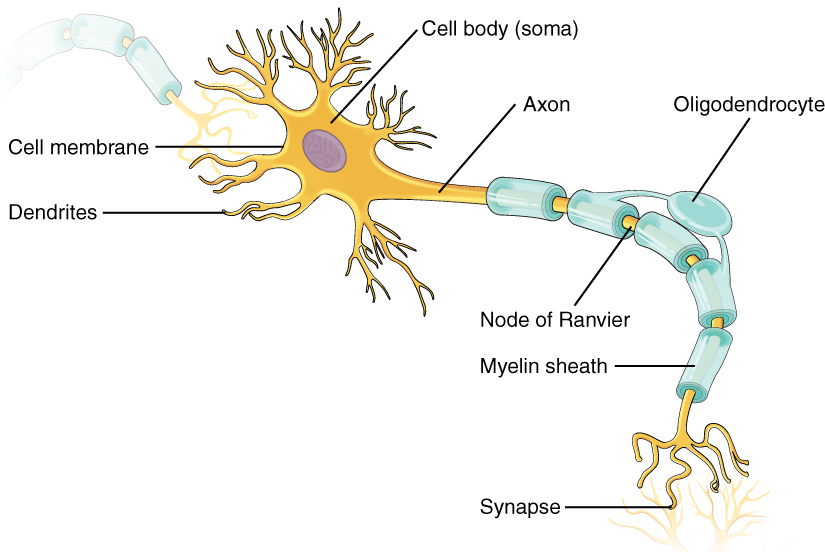
Another view of a “typical” neuron is shown here. As with the previous drawings we’ve looked at, this is a multipolar neuron receiving information from the neuron at upper left and sending information to the neuron at lower right.
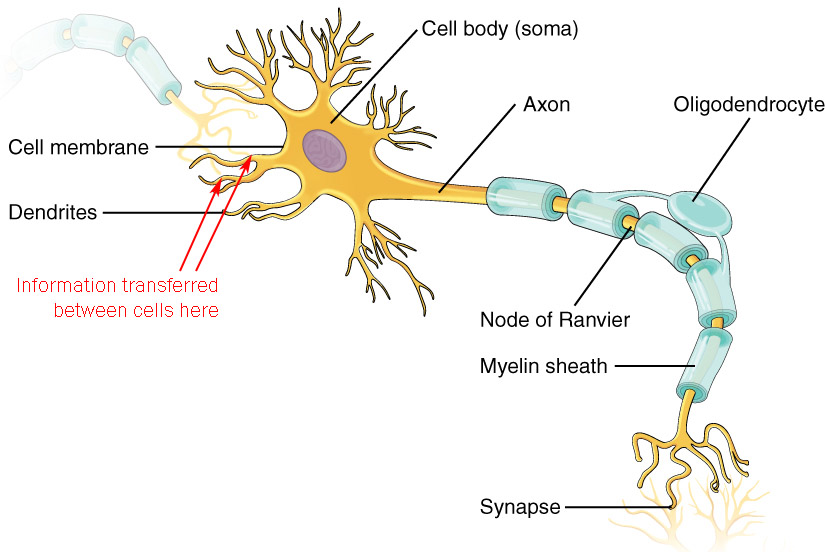
Most, but certainly not all, synapses are between axon terminals and dendrites (the so-called axodendritic synapse). At the axodendritic synapse, the neuron receives information on the dendrites.
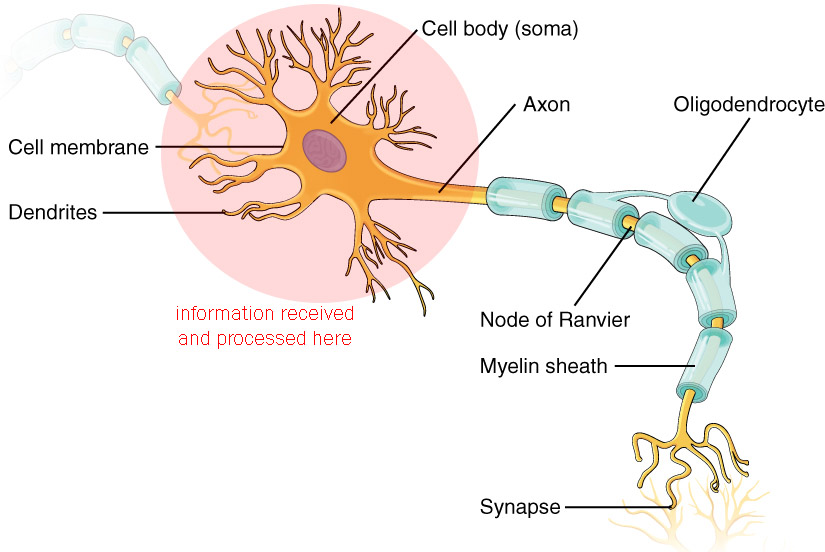
When the information is received, it is processed in the dendritic tree and the cell body (soma), as shown here. The details of this information processing can be quite complex, so the description of how this works will be saved for a later chapter.
The Decision Point
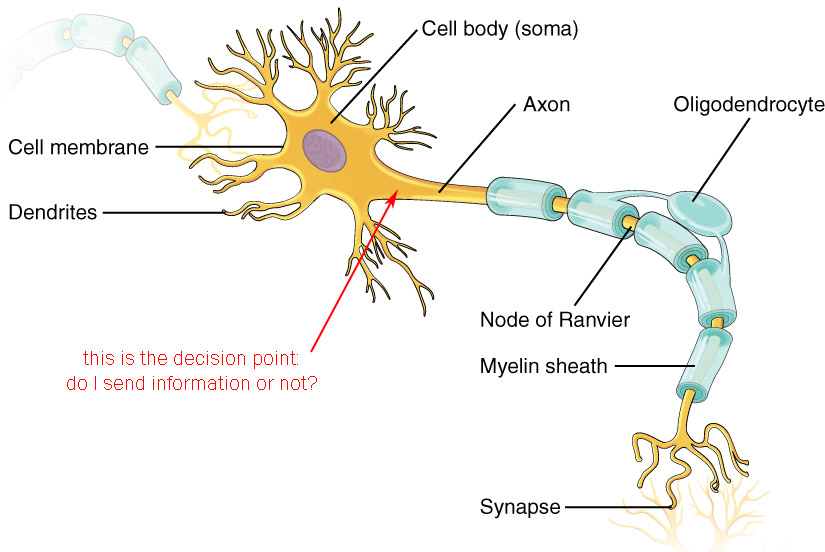
The decision point is where the neuron makes a choice:
- do I fire an action potential or not?
- do I release neurotransmitter or not?
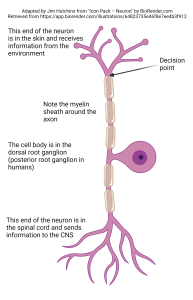
If the neuron is capable of firing an action potential, then it can send its information over a long distance. An example of such a neuron, the unipolar neuron found in the somatosensory system, is shown at left. Nerve endings in the skin gather information about the deflection of the skin, or the release of pain chemicals, or the temperature of the skin. They then make a decision: do I send this information to the central nervous system? Surprisingly, this decision is made within the skin, adjacent to the receptor structure itself. If an action potential is sent, it travels along the axon, by-passing the cell body which is not involved in information processing in this case, and continues into the spinal cord where the action potential will always cause the release of neurotransmitter at the synapse.
If the neuron is not capable of firing an action potential, then it sends its information over a short distance only. These so-called local circuit neurons (sometimes interneurons) have a different decision to make: do I release neurotransmitter? We will see later that a certain voltage must be reached at the synapse before neurotransmitter can be released. In this case, the decision point is the active zone of the synapse where the proteins which support neurotransmitter release are located.
Transporting and Sending Information
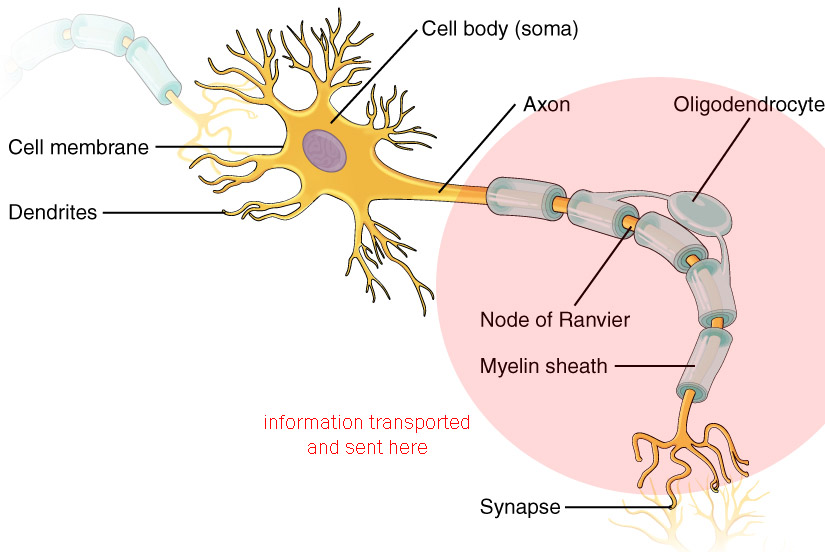
The information, in the form of an action potential, travels from the axon hillock to the axon terminals (never in any other direction, remember) where synaptic release of neurotransmitter occurs.
Synapses
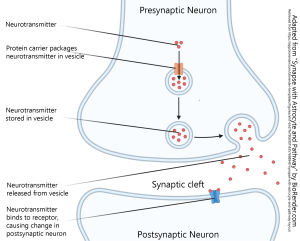
We will read about the details of the chemical synapse and its constituent parts later; there is an entire chapter devoted to this topic. The basics are shown here. The cell sending information is called the presynaptic cell while the cell receiving the information is called the postsynaptic cell. The basic steps are as follows:
- neurotransmitter is packaged into vesicles
- upon arrival of an action potential or other voltage change, the machinery of the synapse causes the release of neurotransmitter
- the neurotransmitter diffuses across the synaptic cleft (enlarged in diagrams for clarity, but generally only 20 nm or about three times the width of the cell membrane
- the neurotransmitter binds to a receptor on the postsynaptic cell and causes an electrical and/or chemical change in the postsynaptic cell
Function Follows Form
As soon as we opened the book, we saw the cover: a famous image of the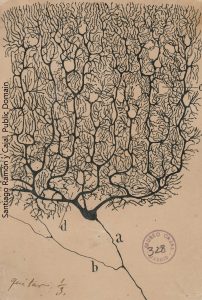 Purkinje cell of the cerebellum drawn by Santiago Ramón y Cajal. Because it’s two-dimensional on your computer screen or textbook page, you don’t appreciate that it really is two-dimensional. The elaborate dendritic tree of the Purkinje cell almost disappears if we rotate it 90° in the plane of the page. That is, it has width and height but not much depth.
Purkinje cell of the cerebellum drawn by Santiago Ramón y Cajal. Because it’s two-dimensional on your computer screen or textbook page, you don’t appreciate that it really is two-dimensional. The elaborate dendritic tree of the Purkinje cell almost disappears if we rotate it 90° in the plane of the page. That is, it has width and height but not much depth.
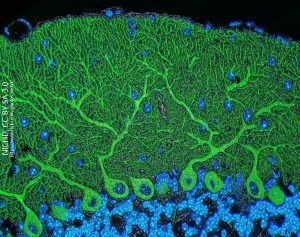
Once the function of the cerebellum was elucidated, many decades after Cajal first drew one, it was clear how the structure of a Purkinje cell relates to its function. We will see later that the cerebellum is responsible for comparing the motor output generated by the primary motor cortex (Brodmann 4; precentral gyrus) to the sensory input coming from the balance and muscle stretch receptors. The motor program is being constantly modified by sensory input as the nervous system adapts to changing conditions or unexpected events.
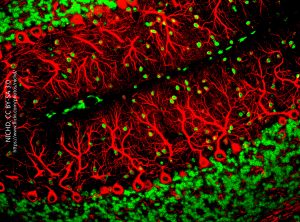 The Purkinje cell must then gather information from tens of thousands of neurons carrying that sensory and motor information; integrate it; and then make a decision whether to fire an action potential or not. If an action potential is generated, it travels a long distance along the single axon (“a” in Cajal’s drawing) to the cerebellar nuclei, where the axon terminal releases the inhibitory neurotransmitter GABA to influence further movement.
The Purkinje cell must then gather information from tens of thousands of neurons carrying that sensory and motor information; integrate it; and then make a decision whether to fire an action potential or not. If an action potential is generated, it travels a long distance along the single axon (“a” in Cajal’s drawing) to the cerebellar nuclei, where the axon terminal releases the inhibitory neurotransmitter GABA to influence further movement.
An Introduction to Cytoarchitectonics
The shape of individual neurons is an important clue to how the brain processes information. But the arrangement of those individual elements is important as well.
Most mammalian cerebral cortex has six layers of neurons visible in a Nissl stain. This six-layered cortex is phylogenetically the newest arrangement of cortex and so is called neocortex for that reason. It was the different size and arrangement of cells in these six layers that led Korbinian Brodmann to differentiate between the different areas that bear his name and numbering system. (All neocortex has six layers, but what’s different is how each of those layers look in a Nissl stain.)
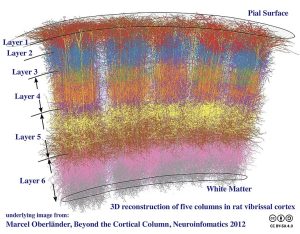

There is another level of organization that runs perpendicular to the layers. Cerebral cortex, including the mouse and rat somatosensory cortex shown here, has an arrangement of neurons so that the set of neurons processing information from a single facial hair (vibrissa) is arranged in a stack called a cortical column. Five cortical columns are shown in this reconstruction. There are extensive interconnections within each column but very little “cross-talk” between columns in this type of cortex. The comparison of columns comes at a later stage of information processing.
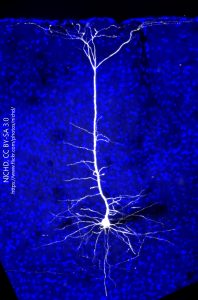 The neurons within columns, as one might expect, have a shape that fits within the column. The white neuron at right with the blue background is a neuron from mouse visual cortex, which also has a strong columnar arrangement. The shape of the cell is called pyramidal, because the cell body is triangular in cross-section and is roughly shaped like an Egyptian pyramid in three dimensions. The cell body lies in layer 5 (layer V). The dendrites that extend out from the cell body stay within the bounds of the column and the single axon seen exiting cortex at the bottom lies in the center of the column. The only dendrites which extend out of the column are seen in the topmost layer of cortex, layer 1 (layer I) which is at the top of the image.
The neurons within columns, as one might expect, have a shape that fits within the column. The white neuron at right with the blue background is a neuron from mouse visual cortex, which also has a strong columnar arrangement. The shape of the cell is called pyramidal, because the cell body is triangular in cross-section and is roughly shaped like an Egyptian pyramid in three dimensions. The cell body lies in layer 5 (layer V). The dendrites that extend out from the cell body stay within the bounds of the column and the single axon seen exiting cortex at the bottom lies in the center of the column. The only dendrites which extend out of the column are seen in the topmost layer of cortex, layer 1 (layer I) which is at the top of the image.
Neocortex (“new cortex”) has six layers. Archicortex (“old cortex”) has three layers. The hippocampus is a good example of archicortex; at the bottom, a single layer of egg-shaped cells is flanked by two layers with few cell bodies and plentiful dendrites and axons.

We can take individual bricks of different sizes and shapes, and stack them carefully so they perform a specific function. This is called architecture.
We can take individual neurons of different sizes and shapes, and stack them carefully so they perform a specific function. This is called cytoarchitecture or cytoarchitectonics. An understanding of both individual neuronal shapes, and the arrangements of neurons, helps us to gain a more complete understanding of the function of nervous tissue.
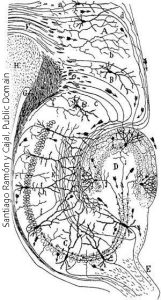
Unlike the columns in visual cortex, which restrict information to a small columnar geographic area of cortex, processing of visual information at early stages in the retina requires extensive cross-talk between adjacent photoreceptors and bipolar cells. This is shown in yet another drawing by Cajal (his work was both extensive and beautiful). Here, we see cells which make synapses in the inner plexiform layer of the lizard retina. The neurons labeled E, G, H, and I have axons and send information out to the brain. These are called ganglion cells. The cells D and F are axonless, and their name (amacrine) means “without long processes” or in other words, “axonless”. As we will see later, these different ganglion cells send different types of information to the brain, and the different types of amacrine cells share information between ganglion cells in a different way.

Media Attributions
- Neuron Shapes © Crookston, Alexa is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Image of a neuron © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license
- Image of a neuron modified © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- Image of a neuron highlight © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- Image of a neuron © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- Unipolar neuron © BioRender adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Image of a neuron information transport and transfer © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- Parts of a synapse © BioRender adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Green purkinje cells © Tom Deerinck and Mark Ellisman, National Center for Microscopy and Imaging Research (NCMIR) is licensed under a CC BY-NC (Attribution NonCommercial) license
- Red purkinje cells © Yinghua Ma and Timothy Vartanian Cornell University, Ithaca, N.Y. is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Cortical Columns © Bodysurfinyon is licensed under a CC BY-SA (Attribution ShareAlike) license
- Colonna di Foca Rome © MM is licensed under a Public Domain license
- Projection Neuron in Mouse Visual Cortex © Massimo Scanziani is licensed under a CC BY-NC (Attribution NonCommercial) license
- Hippocampus Cajal © Santiago Ramón y Cajal is licensed under a Public Domain license
- Architecture © Geo Swan adapted by Jim Hutchins is licensed under a Public Domain license
- Lizard retinal neurons Cajal © Santiago Ramón y Cajal is licensed under a Public Domain license

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}