The Influence of Neuronal Shape on the Electrical Properties of the Neuron
Caleb Bevan
Objective 9: Predict the function of a neuron from your knowledge of neuronal shape and information processing properties.
Now that we know how neurons process information, we can revisit the discussion of neuronal shapes we started earlier in the course. Let’s remember some of the principles that we will use as we analyze neuronal shape.
Neurons tend to distribute proteins where they are needed. Voltage-gated Na+ channels are needed where action potentials are generated and propagated. Therefore, these are found on the neuronal axon.
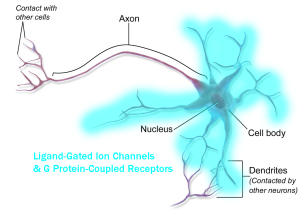
Receptors are distributed in the information-receiving parts of the neuron, generally the dendrites and cell body (although there are many exceptions).
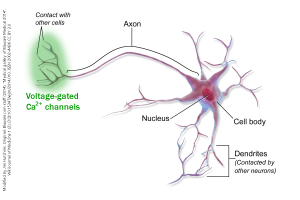
Voltage-gated Ca2+ channels are distributed near the sites of synaptic release, i.e. in the information-sending parts of the neuron, generally the axon terminals (although there are many exceptions).
We need an understanding of the concept of a decision point: this is where a neuron formulates an output. In general, there are two kinds of decision point:
- For firing an action potential, the decision point is the initial segment (axon hillock), where the axon arises from the neuronal cell body (soma). There, we find voltage-gated Na+ and K+ channels, which support the action potential. If enough voltage-gated Na+ channels are opened, the axonal membrane reaches threshold, and an action potential becomes an inevitability. If an action potential reaches the axon terminal, then neurotransmitter release is almost a certainty.
- For neurotransmitter release, the decision point is the active zone of the synapse. There, we find voltage-gated Ca2+ channels, which support the release of neurotransmitter. If enough voltage-gated Ca2+ channels are opened, the active zone reaches threshold, and neurotransmitter release becomes an inevitability.
We need an understanding of the concepts of spatial summation and temporal summation. The shape of a neuron’s dendrites and soma influence the processing that goes on in those structures. In general, the shape and spread of a dendritic tree give us information about where the input is coming from. The position of the dendrites and dendritic spines relative to the decision point give us information about how much influence those inputs have: because of spatial summation, faraway synaptic inputs have little influence on the decision point while nearby synaptic inputs have a large influence.
The length of the axon tells us where the information is being sent. In the retina and some other structures, there are axonless neurons (the word amacrine used to describe some retinal cells means, literally, “axonless”). That tells us that no action potential decision points exist and the processing must be aimed at the decision point at the active zone of synapses. Contrariwise, if a neuron needs to send information over a long distance, it may have very little dendritic branching and/or the soma may not get involved in processing. We see this strategy in the somatosensory (touch) neurons of the skin surface.
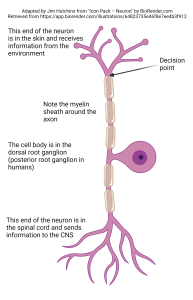
The “job” of this cell is to make a decision about whether to fire an action potential in the skin surface itself. Accordingly, the decision point for firing an action potential is in the skin itself. The modified dendrites in the skin carry out any necessary information processing, and the decision is made. The axon begins where the modified dendrites end. The cell body is found far away, and its job is to provide metabolic support. Unlike most neuronal somata, the cell body of somatosensory neurons does not get involved in information processing. Since there is a single process coming out of the cell body, which branches distally and proximally, this type of neuron is called unipolar.
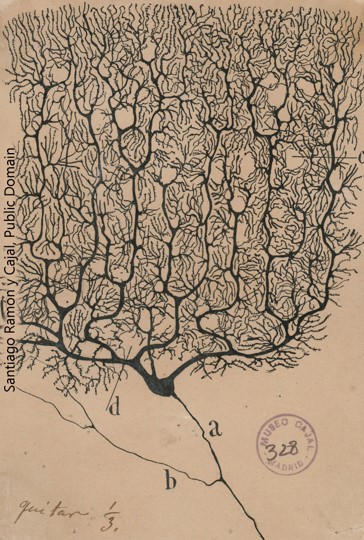
The Purkinje cell of the cerebellum, in contrast, is tasked with picking up information over a wide area from a large number of input cells. This allows the Purkinje cell to make a calculation comparing the expected body movement to what is actually happening. A single axon projects to the so-called “deep” cerebellar nuclei, where the Purkinje cell releases the neurotransmitter GABA to exert an inhibitory influence on the postsynaptic cells it connects to.
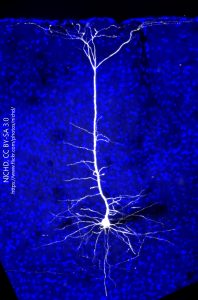
This cell, from the cerebral cortex, is both receiving and sending information from a narrow column of cells. This is typical of cerebral cortex. Note the pyramidal (triangular) shape of the cell body and that some dendrites extend into nearby columns to pick up information from nearby columns. Outside of that cross-talk, the bulk of processing is done in a narrow cylinder which extends perpendicular to the cortical surface, from pia to white matter.
 Most cerebral cortex has six layers, and each layer has a slightly different function. Because it appears fairly recently in the evolution of brains, it is called neocortex. The hippocampus, shown here, has just three layers and is an example of a phylogenetically older type of cortex called archicortex. Note that some cells stay in one of the three layers, while others extend in a columnar fashion through all three layers.
Most cerebral cortex has six layers, and each layer has a slightly different function. Because it appears fairly recently in the evolution of brains, it is called neocortex. The hippocampus, shown here, has just three layers and is an example of a phylogenetically older type of cortex called archicortex. Note that some cells stay in one of the three layers, while others extend in a columnar fashion through all three layers.

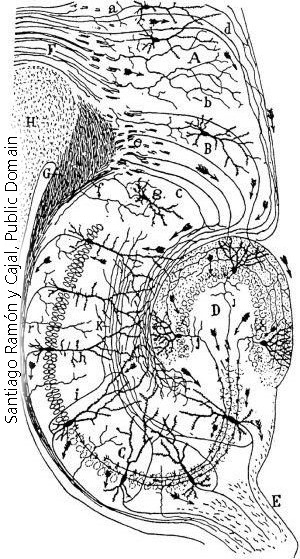
This drawing of the lizard retina by Ramón y Cajal shows some of the different processing strategies within a single cell type. For example, cells E, G, H, and I are all ganglion cells, which send information from the retina to the visual areas of the lizard brain (optic tectum). The retina also has a columnar arrangement, similar to cortex. Cell I is sampling equally from several columns of information flow, while cell H is picking up information from one column at a time, with gaps in between in the dendritic tree. The inner plexiform layer is the fancy name for a region of synapses in the top half of Cajal’s drawing. Different kinds of information are found at different heights in the inner plexiform layer. Cell E is the only ganglion cell that is picking up information at the bottom (innermost) layer of the inner plexiform layer. Between the left-to-right differences in dendritic shape and the top-to-bottom differences, we can surmise that each of these cells is getting a different readout. Cells D and F are the axonless amacrine cells mentioned earlier. Their purpose is to share information between adjacent information columns.
Media Attributions
- GPCR distribution © BruceBlaus adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- Voltage gated calcium channel distribution neuron © BruceBlaus adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- Unipolar neuron © BioRender adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Purkinje cell © Santiago Ramón y Cajal is licensed under a Public Domain license
- Projection Neuron in Mouse Visual Cortex © Massimo Scanziani is licensed under a CC BY-NC (Attribution NonCommercial) license
- Hippocampus Cajal © Santiago Ramón y Cajal is licensed under a Public Domain license
- Lizard retinal neurons Cajal © Santiago Ramón y Cajal is licensed under a Public Domain license

{kind=link}
{kind=link}
{kind=link}
{kind=link}