Gross Anatomy of the Eye
Jim Hutchins
Objective 1: Explain how the gross anatomy of the eye contributes to the initial processing of the visual signal.
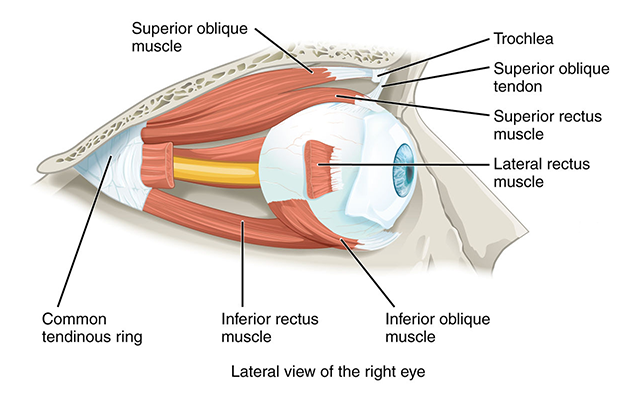 There are six extraocular muscles which are, as the name tells us, outside the eye. Working under the direction of a cortical area called the frontal eye fields, they guide the eyes to an exact, calculated target.
There are six extraocular muscles which are, as the name tells us, outside the eye. Working under the direction of a cortical area called the frontal eye fields, they guide the eyes to an exact, calculated target.
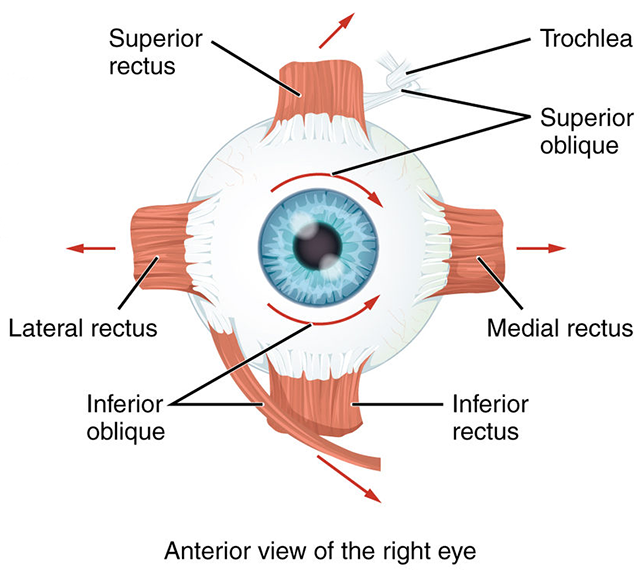 The six extraocular muscles occur in pairs. There is a lateral rectus and a medial rectus. These move each eye along a horizontal plane, from left to right.
The six extraocular muscles occur in pairs. There is a lateral rectus and a medial rectus. These move each eye along a horizontal plane, from left to right.
There is a superior rectus and an inferior rectus. These muscles move each eye along a vertical plane, from up to down.
Finally, there are two oblique muscles which move the eye in more complicated ways that involve rotation and angular movement. These are the superior oblique and inferior oblique.
The cell bodies of lower motor neurons (α motor neurons) which innervate these muscles are found in three cranial nerve nuclei in the brainstem. The largest group forms cranial nerve III (the oculomotor nerve) which innervates the medial rectus, superior rectus, inferior rectus, and inferior oblique. The oculomotor nucleus makes up a large part of the midbrain tegmentum just anterior to the cerebral aqueduct.
The smallest group of neurons innervates the superior oblique muscle. Their axons form cranial nerve IV (the trochlear nerve), which exits the midbrain on the posterior (dorsal) side and wraps around the brainstem. This nerve is named after a bony loop in the eye socket called the trochlea (Latin: “pulley”). The tendon of the superior oblique runs through this loop, which redirects its action as shown in the diagram above.
There is a third group of neurons in the lower pons, near the junction of the pons and medulla. Their axons form the abducens nerve, which pulls the eye away from the midline. The Medical English term for this is “abduct”, so the activation of the abducens nerve abducts the eye.
The eye is divided into three confusingly-named regions: the anterior chamber (between the cornea and iris); the posterior chamber (surrounding the lens); and the vitreous chamber.
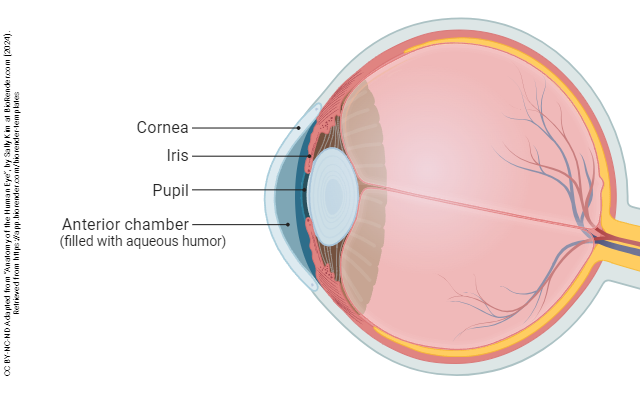
The most anterior part of the eye is a clear window called the cornea. The cornea is responsible for most of the refractory (focusing) power of the eye, as is clear from the use of contact lenses, or from its modification in the surgery called laser-assisted in situ keratomileusis (LASIK).
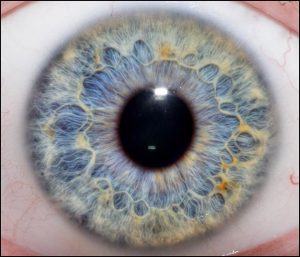
The iris also contains muscular tissue but its job is to change the size of the hole in the center of the iris — the pupil. Smooth muscle fibers which run around the rim of the pupil are called the pupillary sphincter and smooth muscle fibers which run radially in the iris are called the pupillary dilator. The main job of the iris is to shield the photoreceptors from an excessive number of light particle-waves (photons).
When there are very few photons around (i.e. in the dark), the pupillary sphincter relaxes and the pupillary dilator contracts, which increases the size of the pupil to its maximum size.
When there are a lot of photons around (i.e. in bright sunshine), the pupillary sphincter contracts and the pupillary dilator relaxes, reducing the size of the pupil to the smallest possible size and reducing the number of photons that reach the retinal photoreceptors.
A small pupil also helps the eye focus, one of the main reasons it’s easier to see objects more clearly in bright light than in dim light. A pinhole camera is able to focus an image without a lens, which is why it is used to visualize a solar eclipse. As is well known, staring directly at the sun in an eclipse can allow so many photons to flood into the eye, even through a small pupil, that damage to the photoreceptors results.
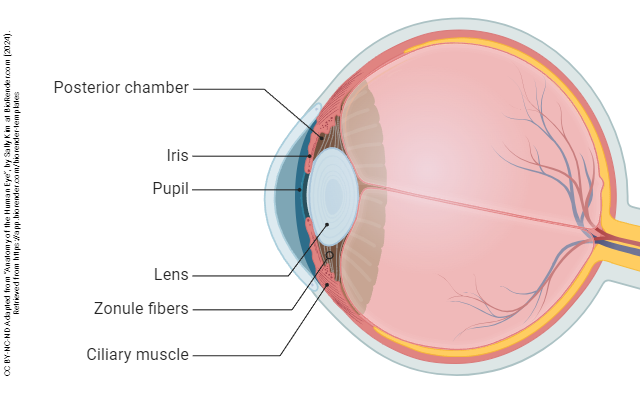
The lens and associated structures are found in the posterior chamber, the space lying between the iris and vitreous body.
The lens also participates in refraction, but unlike the cornea, its refraction is adjustable — at least early in life. A circular muscle called the ciliary muscle, part of the ciliary body, extends rope-like fibers called the suspensory ligaments which attach to the capsule surrounding the lens. When the eye focuses on a far object, the suspensory ligaments pull on the lens, flattening it.
When the eye focuses on a near object, the ciliary muscle relaxes, the suspensory ligaments go slack, and the natural elasticity of the lens pulls it into a rounder shape, allowing the eye to focus on near objects.
The problem occurs as we age and the natural elasticity of the lens decreases. It fails to round up as effectively as it had before, and we are stuck with the ability to only focus on faraway objects. This condition is a natural part of aging and is called presbyopia (literally, “elder vision”).
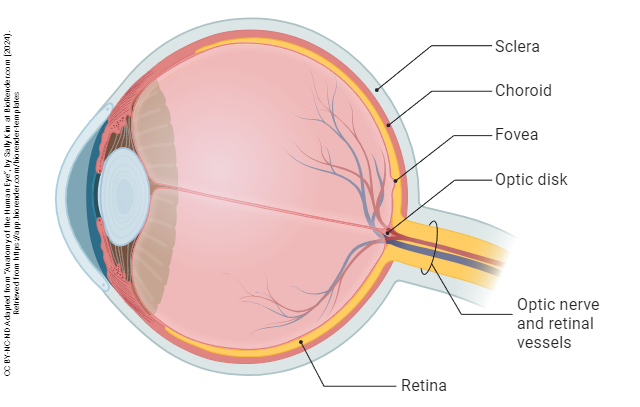
The shape of the eye is maintained by a jelly-like substance called the vitreous humor (“glassy liquid”).
Media Attributions
- Extraocular muscles © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license
- Extraocular muscles © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license
- Anterior chamber © Sally Kim adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Iris © Sarah Cartwright is licensed under a CC BY (Attribution) license
- Posterior chamber © Sally Kim adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Retina and related structures © Sally Kim adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
