The Action Potential in Myelinated Axons
Caleb Bevan
Objective 9: Construct a model of the action potential in myelinated axons.
Traditionally, physiology and neuroscience courses have taught that the mechanisms underlying the action potential are the same in both unmyelinated axons and myelinated axons. The original work by Hogdkin and Huxley in the 1950s, using the squid giant axon, has held up well over the subsequent 75 years. However, mounting evidence has accumulated which suggests that the mechanisms underlying the characteristic action potential waveform are different in unmyelinated and myelinated axons.
The waveform is the same, it’s just what contributes to that waveform is different. 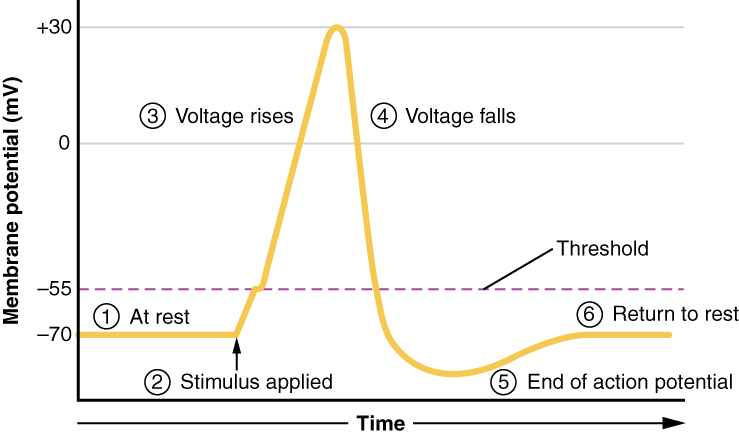
Recall that at the start of the action potential, the inside of the cell is negatively charged, and voltage-gated sodium channels begin to be activated by a positive voltage deflection to threshold at the axon hillock (initial segment). We called this the decision point because if the voltage reaches threshold (the decision), then enough voltage-gated sodium channels open to cause further upward (more positive, depolarizing) deflection of the membrane voltage. This is the phase marked “2” here.
The inside of the neuron is negatively charged and voltage-gated sodium channels are open. It looks like this:
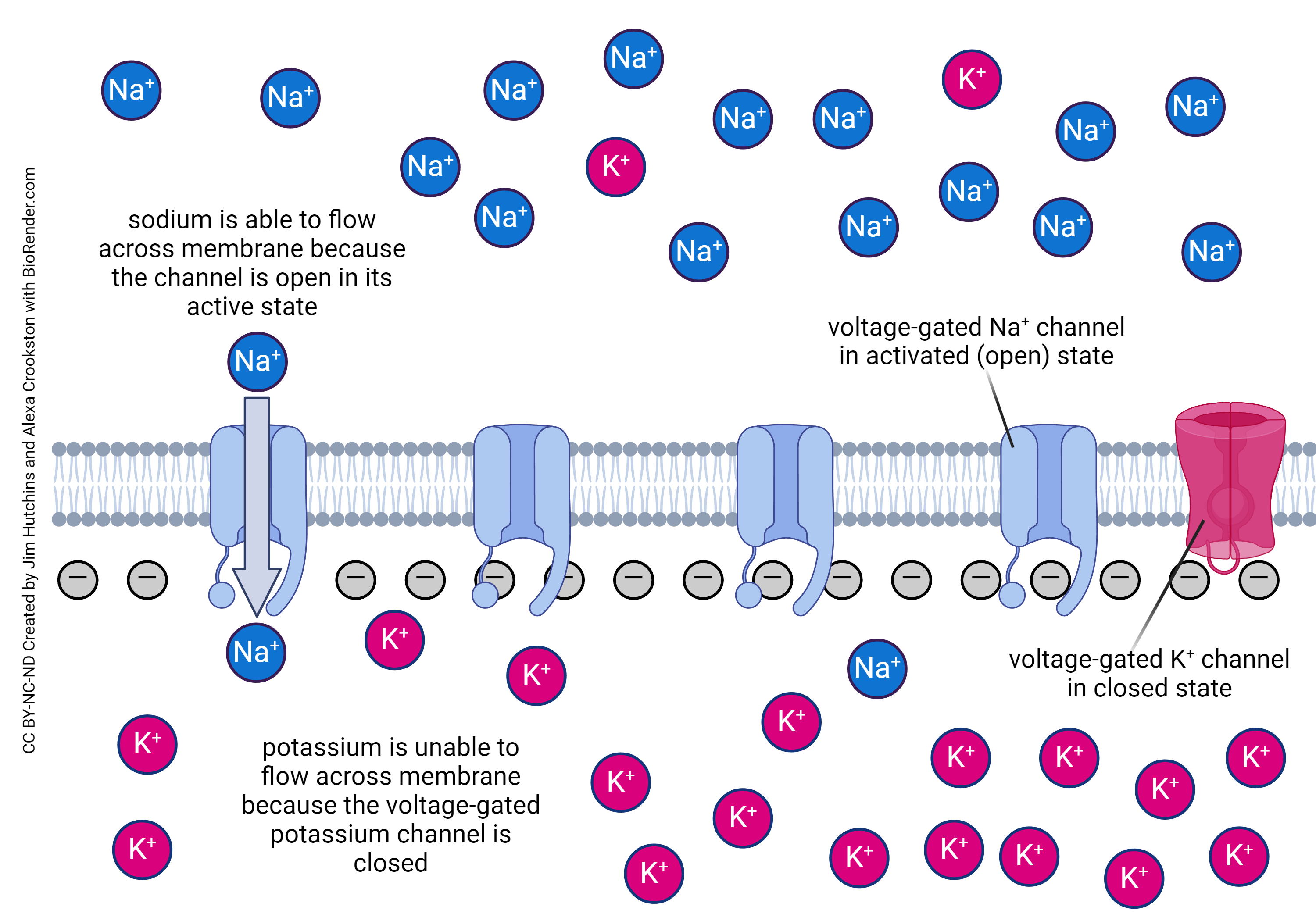
The explosive rise in membrane potential as positively charged Na+ ions rush in through voltage-gated Na+ channels and the neuron depolarizes, happens during the phase marked “3” here.
What is different in myelinated axons is phase “4”, where the voltage falls.
Recall from Objective 8 that the rising phase of the action potential occurs due to the opening of voltage-gated sodium channels. In unmyelinated axons, like the squid giant axon studied by Hodgkin and Huxley, the falling phase of the action potential is due to the action of slightly slower voltage-gated potassium channels.
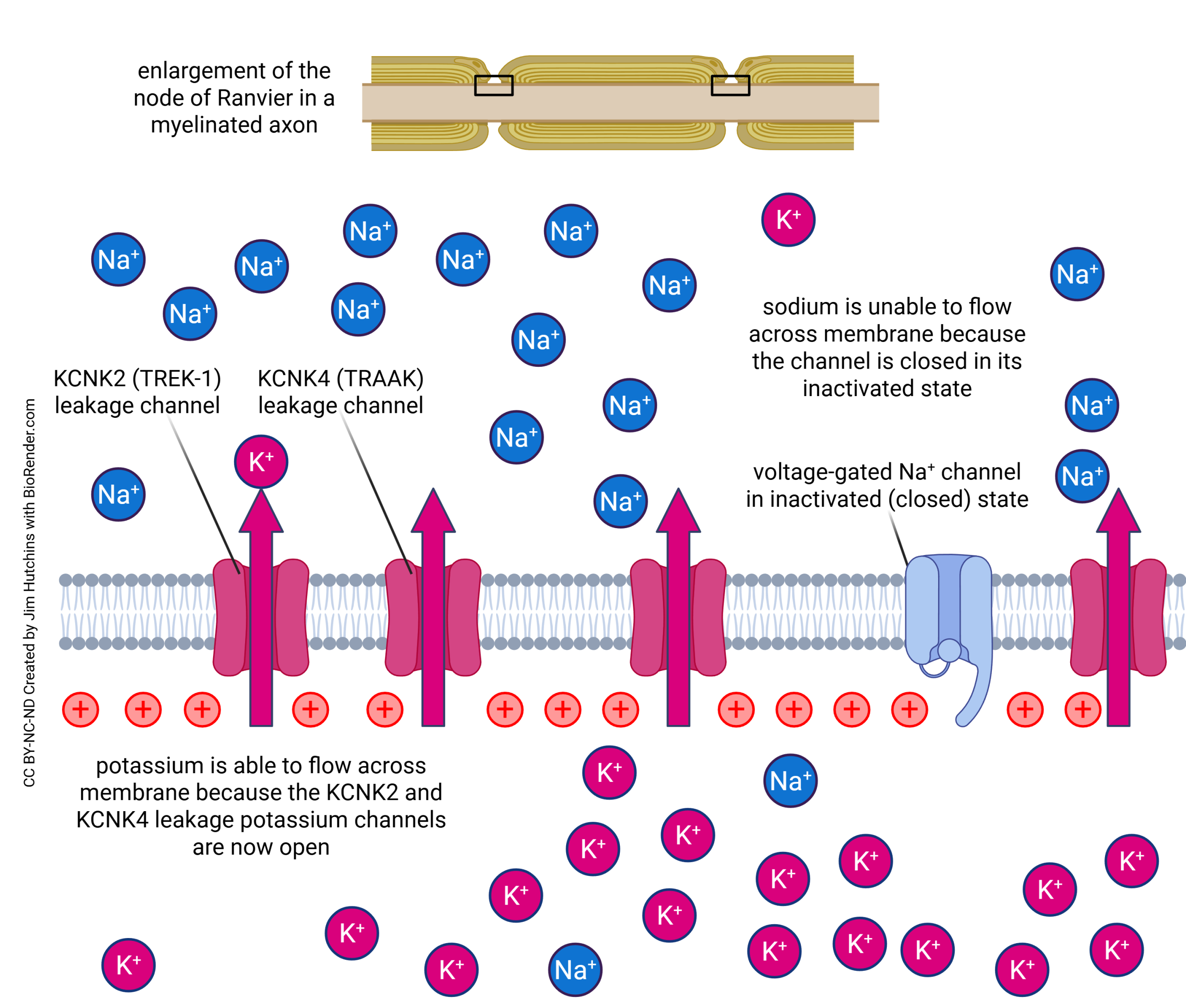
But in myelinated axons, more recent work has shown conclusively that while the rising phase is still due to voltage-gated sodium channels, as described here and in Objective 8, that the falling phase occurs because of a high density of leakage potassium channels. The genes that encode for these channels are KCNK2, which is also called TREK-1, and KCNK4, which is also called TRAAK.
Media Attributions
- Action potential steps © J. Gordon Betts, Kelly A. Young, James A. Wise, Eddie Johnson, Brandon Poe, Dean H. Kruse, Oksana Korol, Jody E. Johnson, Mark Womble, & Peter DeSaix is licensed under a CC BY (Attribution) license
- Rising phase of action potential © Jim Hutchins and Alexa Crookston is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Falling phase of action potential in myelinated axons © Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
