The Inositol Triphosphate / Diacylglycerol / Calcium Second Messenger System
Jim Hutchins
Chapter under construction. This is the first draft. If you have questions, or want to help in the writing or editing process, please contact hutchins.jim@gmail.com.
The cell membrane is made up of phospholipids, molecules with a polar (water-loving, hydrophilic) head group and two non-polar (water-hating, hydrophobic) tails. This is shown in almost every representation of the neuronal cell membrane in this book.
One of these molecules, phosphatidylinositol (PI), plays a key role in intracellular chemical signaling. While the lipid tails vary (these are shown in this skeletal chemical diagram by the zigzag lines), the bridging three-carbon glycerol molecule is invariant.
(The glycerol molecule is shown here in the middle of this diagram with carbons numbered 1, 2, and 3.)
The six-sided figure at the bottom that looks vaguely like a chair is the sugar inositol. Its six carbons are numbered 1-prime (1′), 2-prime (2′), and so forth. There is a phosphate group (PO4) attached to both the glycerol and inositol in a bond called a phosphodiester linkage. That bridging phosphate is connected to the 1′ carbon. The 4′ and 5′ carbons of inositol are also bound to phosphate groups in the molecule shown here, phosphatidylinositol 4′,5′-bisphosphate (PIP2). Based on the concentration of PIP2 in red blood cells, scientists estimate that PIP2 makes up about 1% of the phospholipid molecules in the neuronal membrane.
PIP2 has an almost uncountable number of roles in the neuron. These many roles can be sorted into two groups, however.
First, PIP2 interacts directly with ion channels embedded in the cell membrane. An example of this is the interaction between PIP2 and the G protein-gated inwardly rectifying K+ (GIRK) channels mentioned previously. PIP2 can direct components of the neuronal membrane to various locations in the cell. Probably the most important direct action of PIP2, however, is the interaction with the neuronal cytoskeleton. PIP2 stabilizes actin filaments and is essential for the growth of neurons both in developing and in mature neurons.
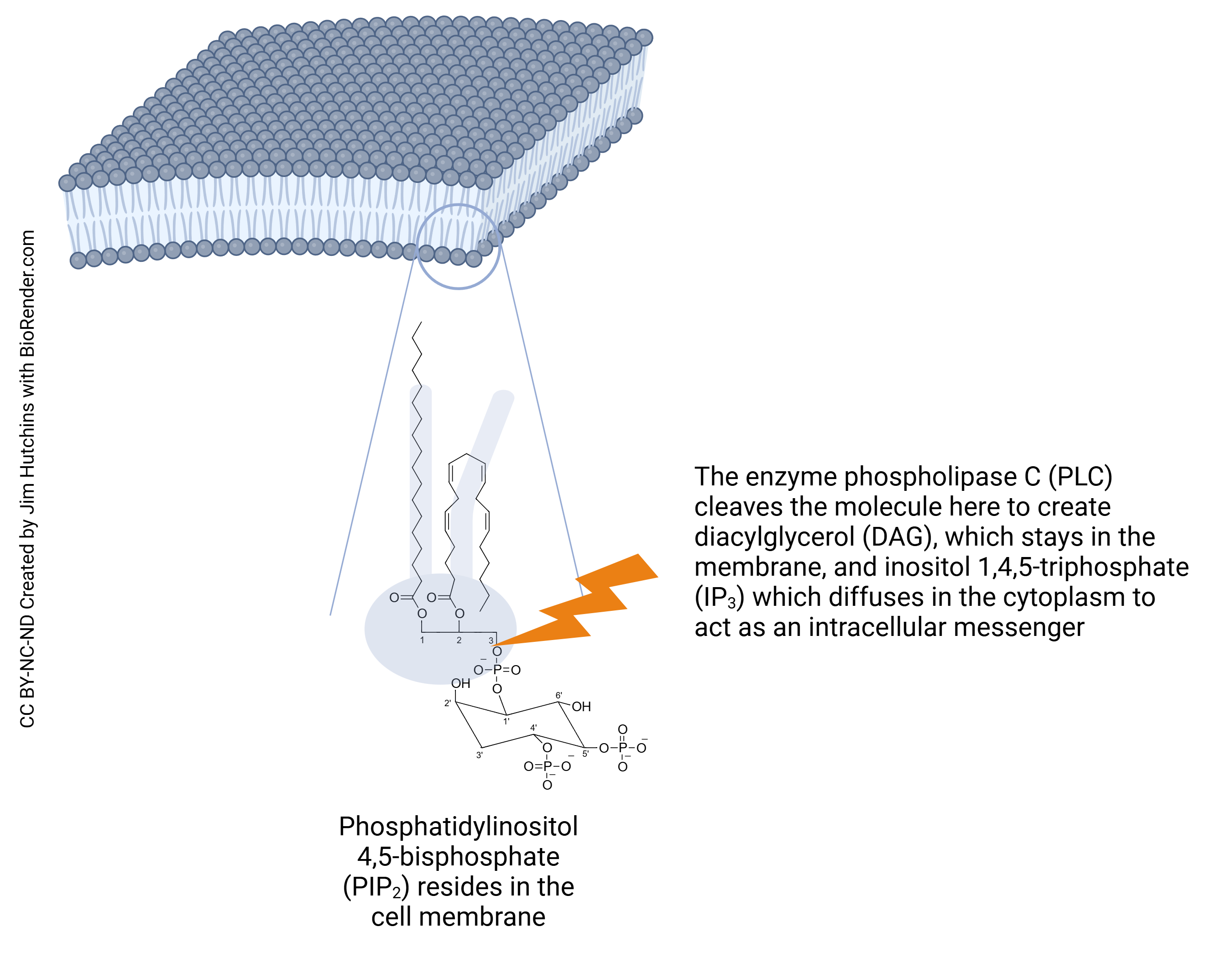
The second group of functions dependent on PIP2 use this molecule as a substrate for the creation of other intracellular signaling molecules. The enzyme phospholipase C cleaves the PIP2 molecule between the #3 carbon of glycerol and the phosphate group, creating two new signaling molecules: diacylglycerol (DAG), which stays anchored in the neuronal membrane, and inositol 1, 4, 5-trisphosphate (IP3), which moves around in the cytoplasm as a second messenger molecule.
DAG activates a number of other systems in the neuron:
- protein kinase C (PKC), which in turn triggers intracellular signals and gene expression leading to proliferation, survival, apoptosis, and motility;
- protein kinase D (PKD), which controls proliferation, survival, gene expression, vesicular trafficking, and neuronal development;
- DAG kinases (DGKs), involved in proliferation and the cytoskeleton;
- chimaerin, which is also involved in neuronal development;
- Munc13, one of the key proteins in vesicle fusion and exocytosis.
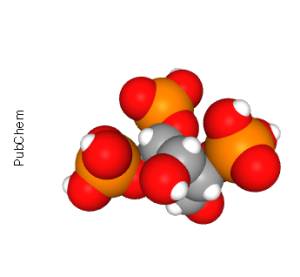
The second signaling molecule created from the enzymatic cleavage of PIP2 is the sugar-phosphate part, inositol 1,4,5-triphosphate (IP3). The main role of IP3 is as a ligand for the ryanodine receptor, which releases Ca2+ from intracellular stores (generally, the smooth endoplasmic reticulum in neurons and the sarcoplasmic reticulum in muscle cells).
When these systems are triggered by a G protein-coupled receptor, the G protein which transduces the signal is called a Gq.
(Remember that we have already seen Gs proteins which stimulate cAMP production, and Gi proteins which inhibit cAMP production. Unlike the letters “s” and “i”, the letter “q” in Gq proteins carries no meaning at all and is merely a convenient letter of the alphabet.)
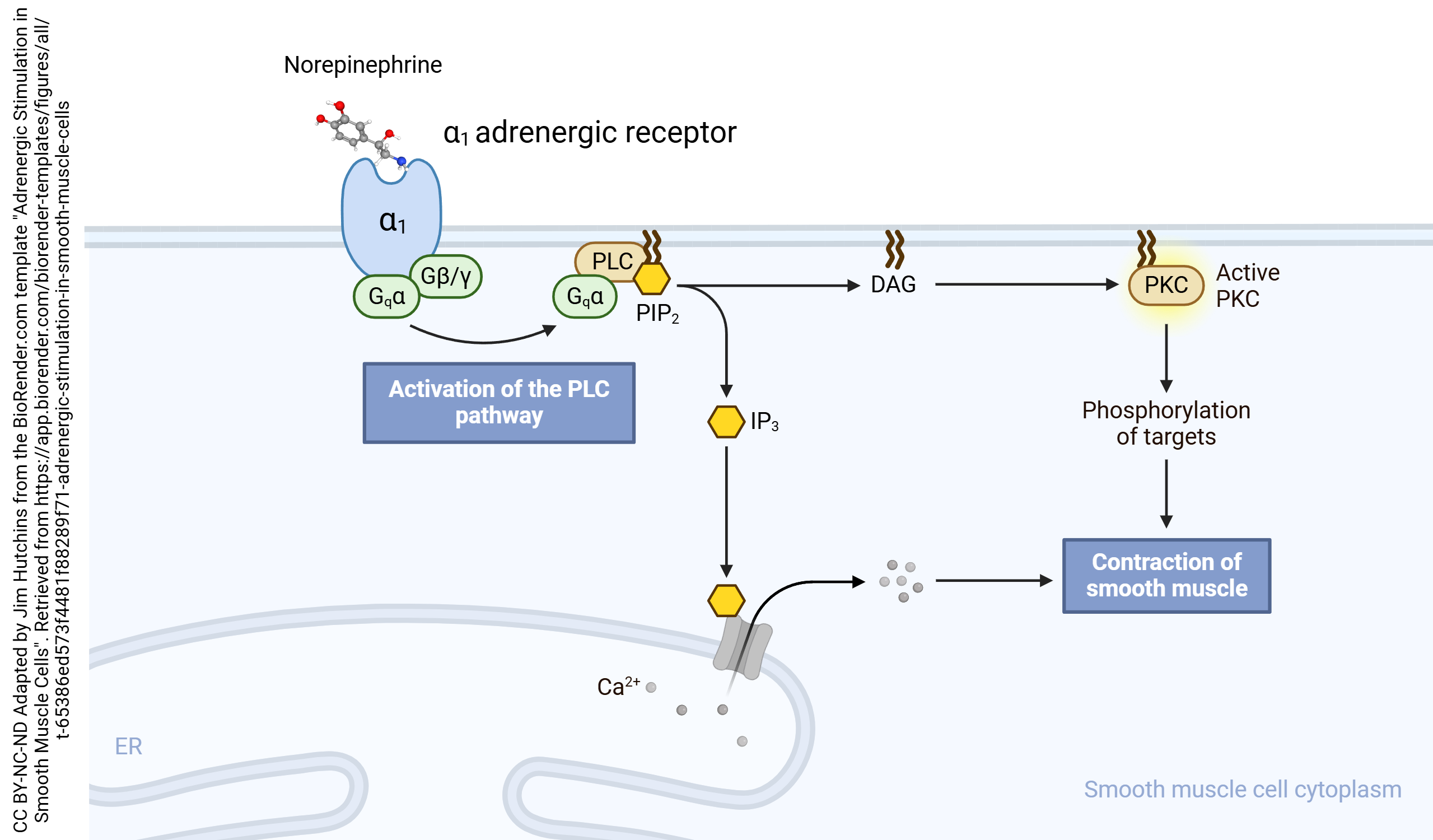 An example of a receptor whose signal transduction is dependent on a Gq protein is the alpha-1 (α1) adrenergic receptor. (Even though the naturally occurring ligand for this receptor is either called norepinephrine or noradrenaline, the receptor is called adrenergic.) The αq subunit carries a lipid anchor which holds it in the plane of the membrane. It slides in a plane just inside the inner leaflet of the neuronal membrane where it collides with, and activates, phospholipase C. PLC splits PIP2 into DAG, which activates protein kinase C, and IP3, which releases Ca2+ from intracellular stores.
An example of a receptor whose signal transduction is dependent on a Gq protein is the alpha-1 (α1) adrenergic receptor. (Even though the naturally occurring ligand for this receptor is either called norepinephrine or noradrenaline, the receptor is called adrenergic.) The αq subunit carries a lipid anchor which holds it in the plane of the membrane. It slides in a plane just inside the inner leaflet of the neuronal membrane where it collides with, and activates, phospholipase C. PLC splits PIP2 into DAG, which activates protein kinase C, and IP3, which releases Ca2+ from intracellular stores.
Media Attributions
- PIP2, DAG, and IP3 © Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- d-Myo-inositol-1-4-5-triphosphate Conformer © PubChem adapted by Jim Hutchins is licensed under a Public Domain license
- Adrenergic Stimulation in Smooth Muscle Cells © BioRender adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- GPCR and Akt_mTOR signaling © BioRender adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- Growth factor receptors and Akt_mTOR signaling © BioRender adapted by Jim Hutchins is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
