Synaptic Pruning in the Retinogeniculate System
Jim Hutchins
Chapter under construction. This is the first draft. If you have questions, or want to help in the writing or editing process, please contact hutchins.jim@gmail.com.
Synaptic Pruning in the Retinogeniculate Pathway
Processing in the retina converges on the output cell of the eye, the retinal ganglion cell. Retinal ganglion cell axons bundle together and grow to a target in the thalamus called the lateral geniculate nucleus (LGN). In all mammals, the LGN contains a retinotopic map of the visual world. That is, if two points are adjacent in visual space (say, your index and middle finger held out at arm’s length), then they are represented by adjacent retinal ganglion cells, adjacent LGN relay cells, and adjacent visual cortical cells.
There are two ways that synaptic pruning refines the pathway between eye and brain.
First, as the neuroscientist Carla Shatz famously said, “cells that fire together, wire together”; synaptic pruning makes sure that only cells that are active about the same time are marked as adjacent. Think of the wave in a large stadium. You would know who your neighbor is by the fact they stand up at almost the same time you do, even if you never perceived the larger pattern in the entire stadium. These nearest-neighbor relationships are maintained by pruning non-neighboring synapses.

Second, in primates and closely related mammals, the LGN has six layers of cell bodies, numbered from deep to superficial. In the tree shrew, layers 2, 3, 4, and 6 receive synapses from the opposite-side (contralateral) eye, while layers 1 and 5 receive synapses from the same-side (ipsilateral) eye. This relationship, too, is maintained by synaptic pruning, so that inappropriate synapses are removed early in development.
Years ago, Vivien Casagrande and Jim Hutchins used the tree shrew as a model system. Tree shrews are native to the forests of southeast Asia. 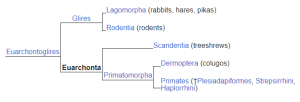 When European naturalists first encountered them, they were mistaken for squirrels. Later, the paleoanthropologist Wilfrid Le Gros Clark classified them as prosimian primates based on their brain structure. In particular, the LGN of the tree shrew looks amazingly like the primate LGN. More recent evidence indicates that tree shrews, flying lemurs, and primates fit within a grandorder called Euarchonta (“true ancestors”). One possible arrangement of tree shrews, flying lemurs, and primates within this grandorder is shown.
When European naturalists first encountered them, they were mistaken for squirrels. Later, the paleoanthropologist Wilfrid Le Gros Clark classified them as prosimian primates based on their brain structure. In particular, the LGN of the tree shrew looks amazingly like the primate LGN. More recent evidence indicates that tree shrews, flying lemurs, and primates fit within a grandorder called Euarchonta (“true ancestors”). One possible arrangement of tree shrews, flying lemurs, and primates within this grandorder is shown.
Tree shrews are altricial; that is, their visual systems are slow to develop and much of visual system development occurs outside the womb. Being both primate-like and altricial made them a perfect model organism for studies of visual system development.
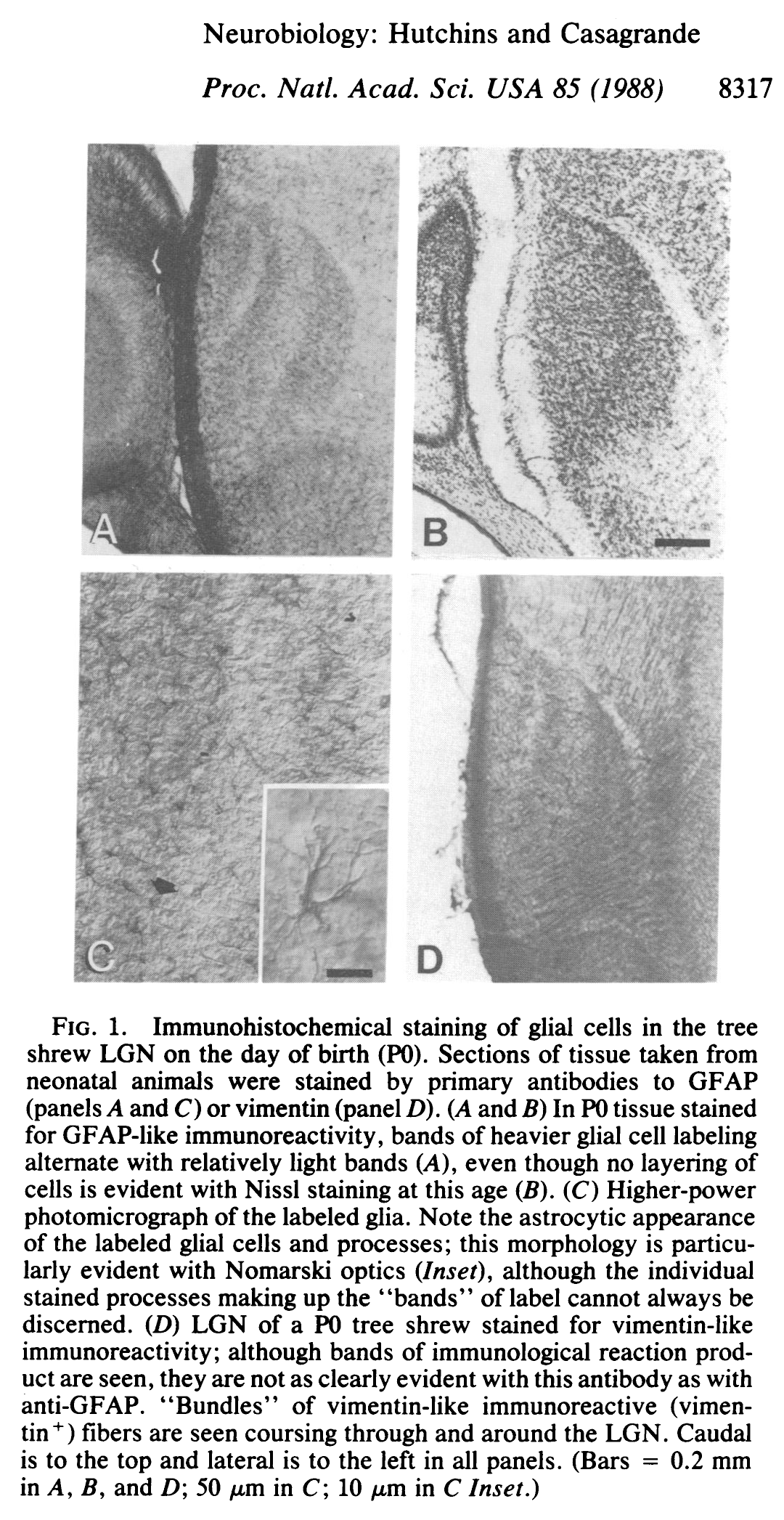
At birth (postnatal day zero, or P0), staining for astrocytes (glial fibrillary acidic protein, or GFAP) reveals a layering of the LGN which is not seen until at least a week later in the cytoarchitecture of neurons.
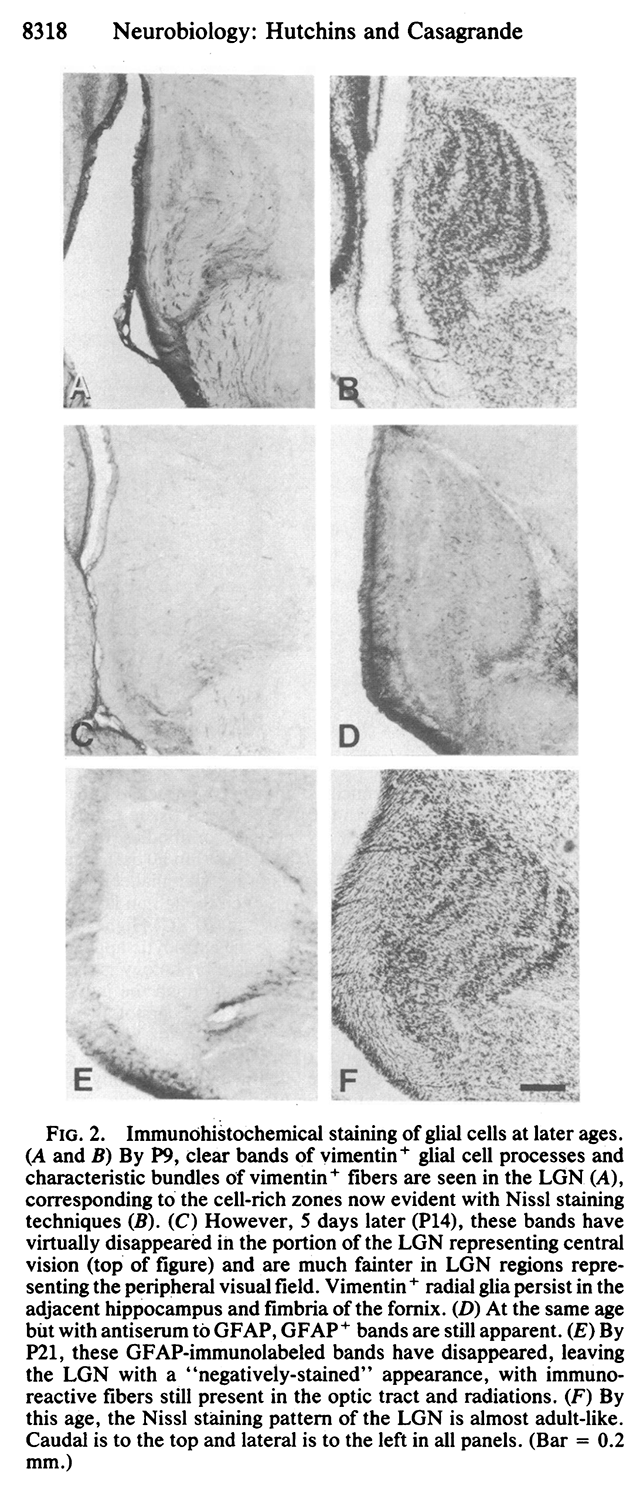
That is, by postnatal day 9 (P9), there are six distinct layers of neurons with cell-free synaptic zones in between (figure 2B below). This resembles the adult pattern of cytoarchitecture (figure 2F).
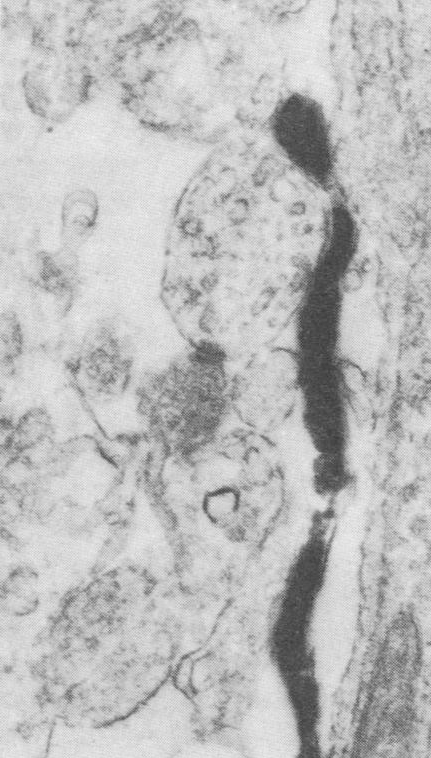
It was not at all clear to the authors what the glial cells were doing there. In retrospect, the association between astrocytes and developing synaptic contacts, seen by immunocytochemistry under the electron microscope, provides us with a clue if we look at it through the lens of what has been discovered. The dark, fuzzy pair of parallel lines in the center of this image is a developing synaptic contact. The glial cell is in close association with this developing synapse. What is the glial cell doing there? Why do glial cells form a layered structure before the Nissl-stained cell bodies of neurons do?
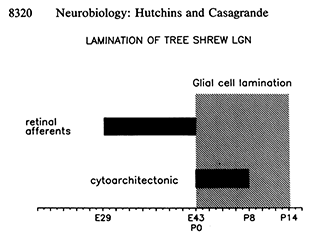 Hutchins and Casagrande offered the timeline at right to understand the role of glial cells in the layering of the tree shrew LGN. Development of the tree shrew is timed from fertilization, termed embryonic day zero (E0). Tree shrews are born after 43 days gestation (E43 = P0). At that time, even though the eyes will not open until about P21 and the visual system is still actively developing, the glial cells are reflecting the location of synaptic pruning.
Hutchins and Casagrande offered the timeline at right to understand the role of glial cells in the layering of the tree shrew LGN. Development of the tree shrew is timed from fertilization, termed embryonic day zero (E0). Tree shrews are born after 43 days gestation (E43 = P0). At that time, even though the eyes will not open until about P21 and the visual system is still actively developing, the glial cells are reflecting the location of synaptic pruning.
Media Attributions
- Tupaia © to.wi is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Tree shrew grandorder euarchonta © Unknown is licensed under a CC0 (Creative Commons Zero) license
- hutchins and casagrande 1988 fig 1 © Jim Hutchins & Vivien Casagrande is licensed under a All Rights Reserved license
- hutchins and casagrande 1988 fig 2 © Jim Hutchins & Vivien Casagrande is licensed under a All Rights Reserved license
- hutchins casagrande 1988 glial cell lamination lgn fig 3c © Jim Hutchins & Vivien Casagrande is licensed under a All Rights Reserved license
- hutchins casagrande 1988 glial cell lamination fig 4 320 px © Jim Hutchins & Vivien Casagrande is licensed under a All Rights Reserved license
