Molecular Properties of the Neuronal Membrane
Abbigail Higgins and Jim Hutchins
Chapter under construction. This is the first draft. If you have questions, or want to help in the writing or editing process, please contact hutchins.jim@gmail.com.
The neuronal membrane is composed mostly of a phospholipid bi-layer with various types of proteins integrated within and around it. Phospholipids are able to form very selective membranes due to their exceptional molecular structure. They have a phosphate group creating the polar hydrophobic head which is then joined to two fatty acid tails. Because of this amphipathic structure, phospholipids will naturally form into a membrane as the hydrophobic tail regions repel the water around and create weak intermolecular attractions between the lipid tails (called van der Waals forces). These fatty acid tails can be a variety of lengths and saturations, which can alter the fluidity of the membrane. The number of proteins and the presence of cholesterol will also alter membrane fluidity. Temperature is also a large factor in membrane fluidity. All healthy cells in the body will manage their membrane fluidity by altering the fatty acid and protein content along the membrane. This process maintains a consistent membrane permeability among different cells.
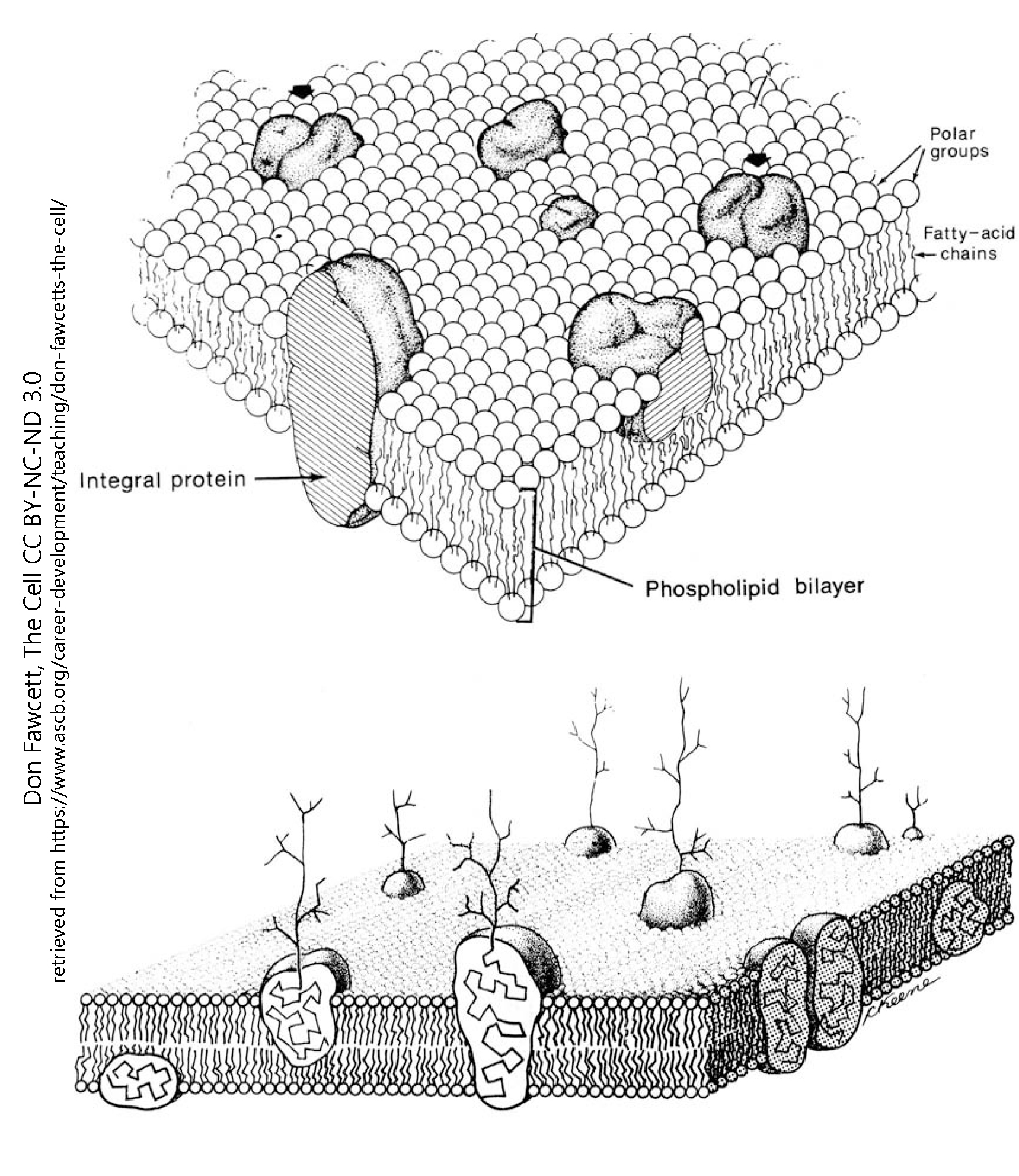
Along the membrane there are a variety of different proteins. They fall into two main categories; peripheral (or surface) proteins, which do not pass through the full membrane and are generally linked to a specific point in the cytoskeleton, and integral proteins, which span the entire membrane and are usually able to shift within the membrane. Some of these proteins are formed into what is known as the cytoskeleton, which gives additional structure and shape to the membrane and serves as an anchoring point for many other proteins. There are many different forms of each type of protein, which will be briefly summarized bellow, then discussed in depth in further chapters.
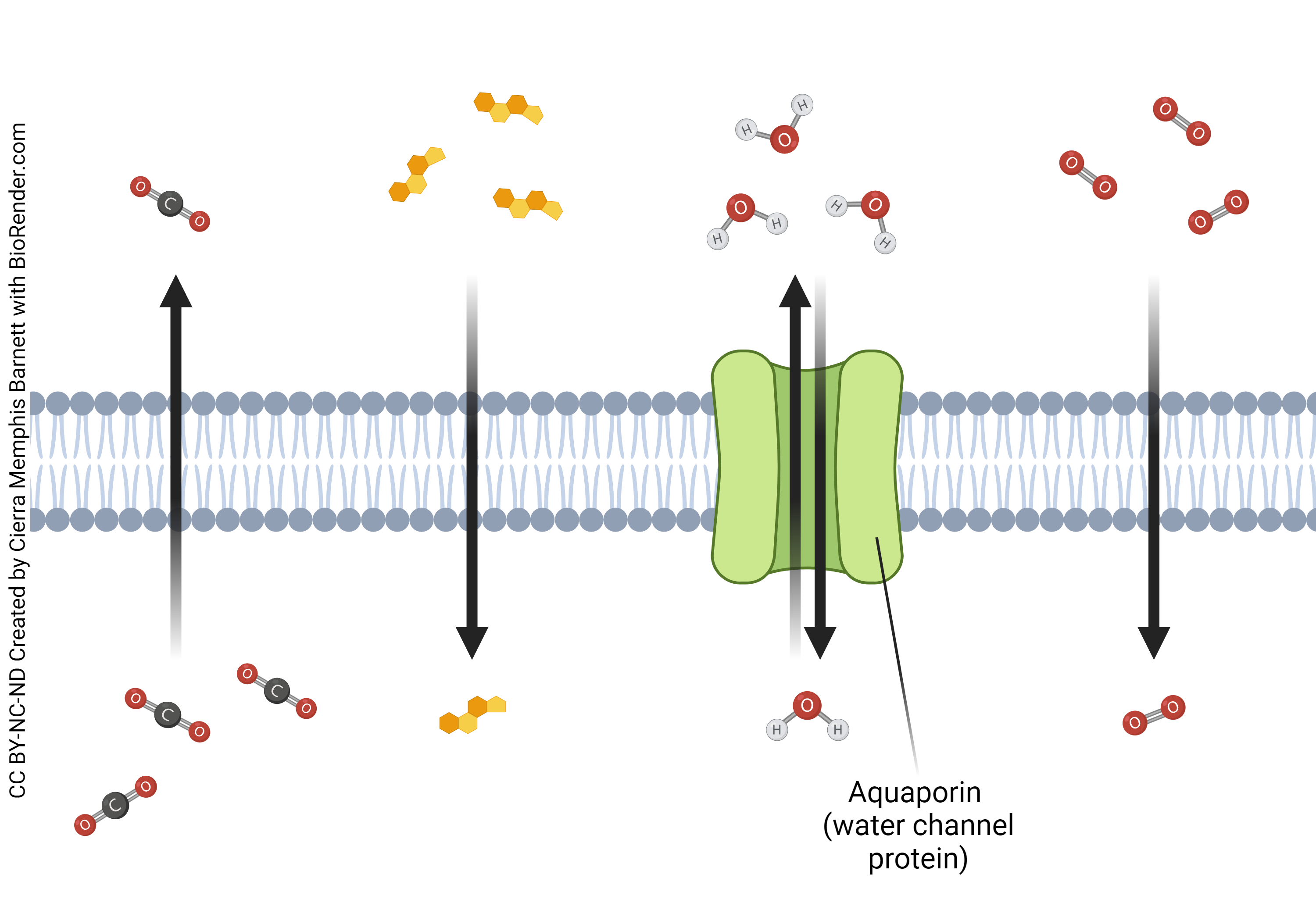 Permeability of the bare phospholipid membrane with no proteins
Permeability of the bare phospholipid membrane with no proteins
The molecular properties of the phospholipid bilayer results in extreme selectivity of the molecules that may pass through. Only small lipid-soluble molecules can break through and pass with simple diffusion (directly through the membrane). Steroid hormones and gasses such as CO2 (carbon dioxide) and N2O (nitrous oxide) are such molecules that are able to pass through the membrane. Water will also at times pass through the membrane, though is able to pass much easier through aquaporins. Larger molecules (such as sugars and proteins) and charged molecules (such as ions) are unable to pass through.
Transporters
Transporter is the broad category which describes all integral proteins that transport molecules across the membrane. This can be none uniport (one type of molecule in one direction), antiport (two types of molecules in opposite directions), or symport (two types of molecules in the same direction). Transporters may use passive or active transport, depending on if they are pushing molecules toward their concentration gradient (from high to low concentrations) or against their concentration gradient (from low to high) respectively. Passive transport, also known as facilitated diffusion, does not require the input of energy because molecules will naturally flow from areas of high concentration to areas of low concentration. Ion channels use this form of transport. Active transport, which is used by pumps, however does require the input of energy. This is usually done in one of two ways. The first is primary active transport, where the transporter uses ATP to power its action. The next is secondary active transport, which uses another reaction to generate energy. This is often through using one molecule passing in its concentration gradient to power another molecule going opposite its concentration gradient.
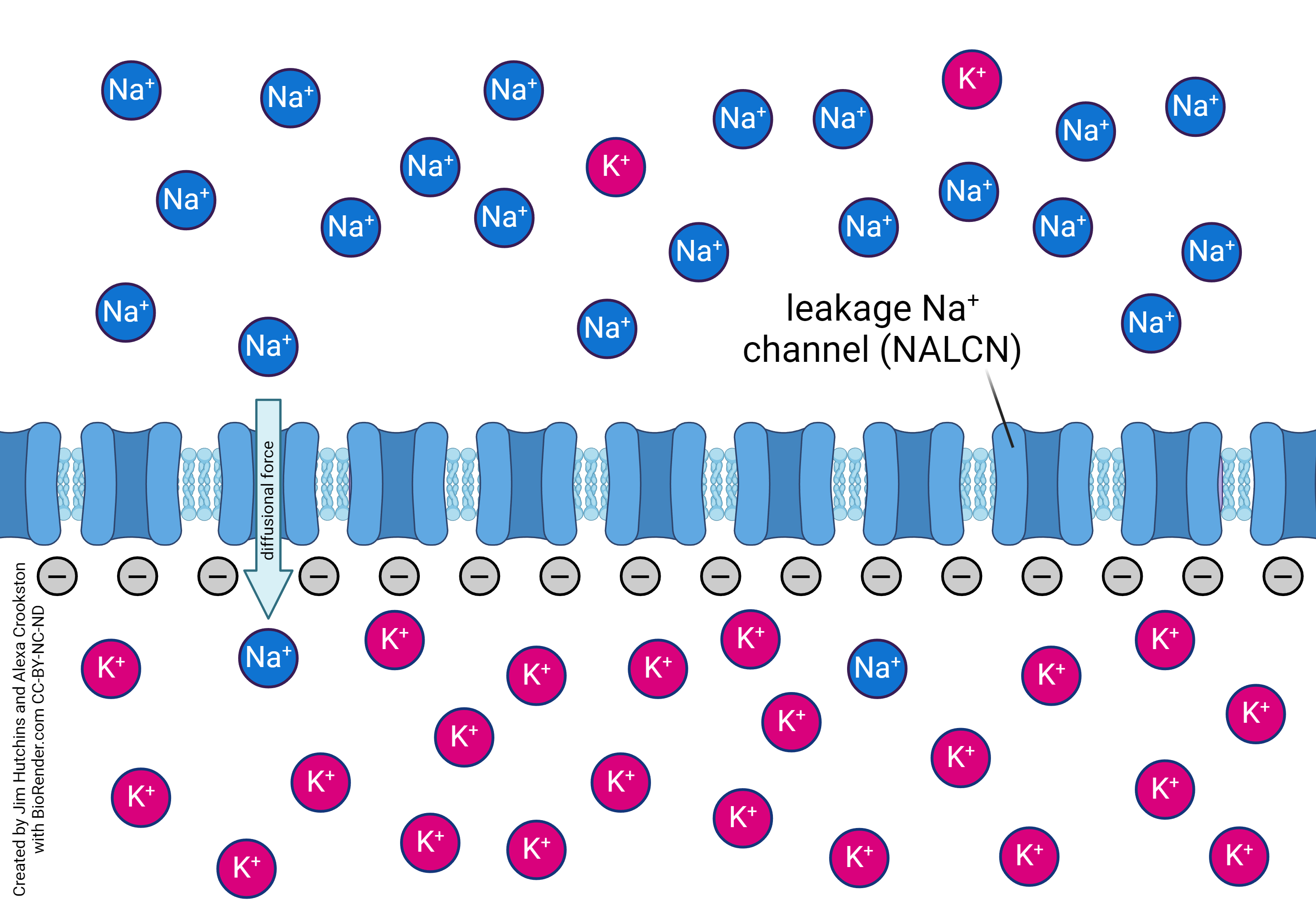
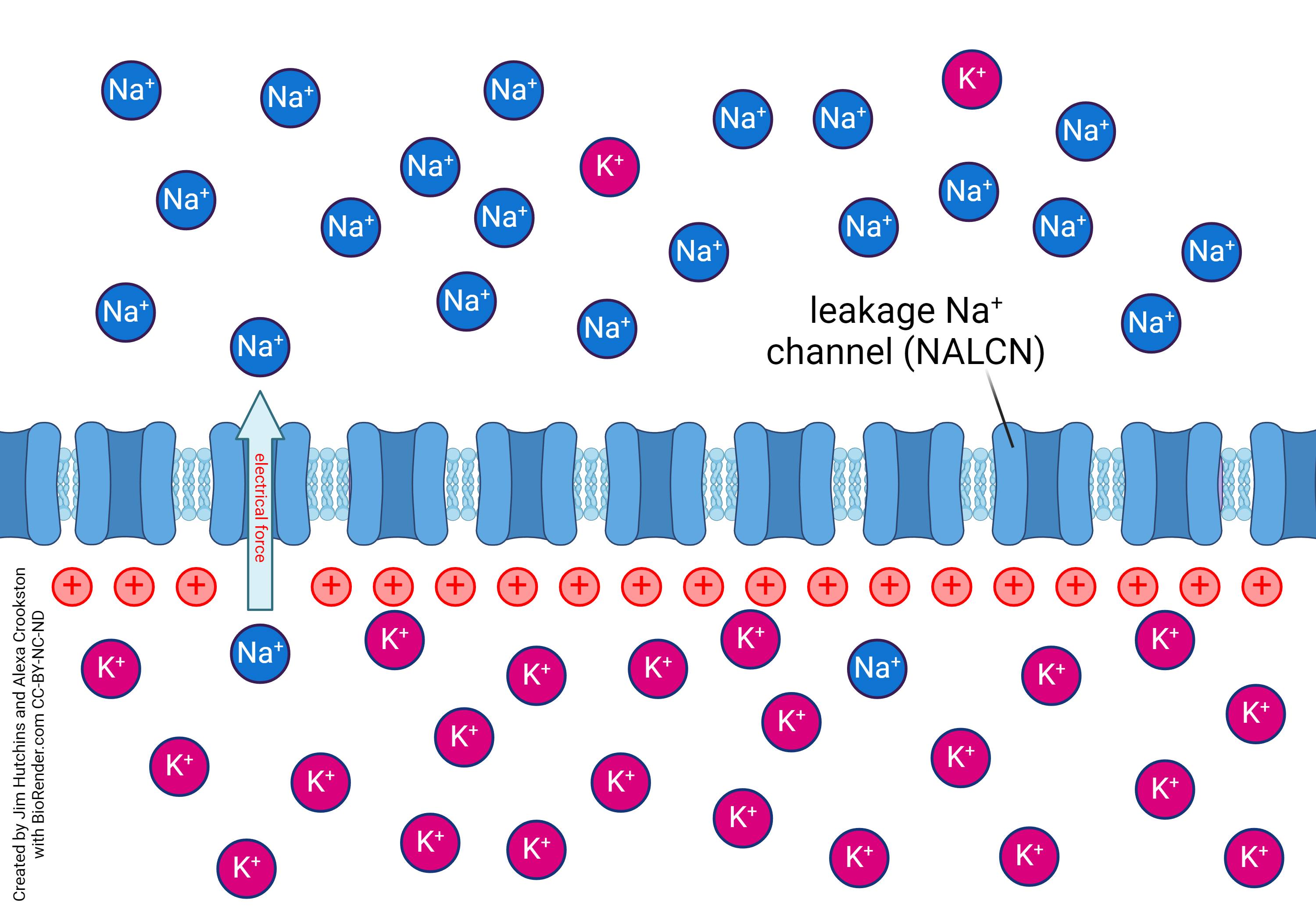
[fix those two images above so that they can sit side by side and still be readable]
Ion Channels
Ion channels are an integral protein which allows charged molecules (ions) to move with their concentration gradient (passive transport) through the cell membrane. This concentration gradient is mostly determined by concentration of ions inside and outside of the cell, but is also effected by the differing charge inside and outside of the cell. Sodium, potassium, calcium, and chloride are the most common and important ions used by the body, that will pass through their respective channels. Each channel will have a specific ion that they will move, so there are many different ion channels throughout the cell. There are four main types of ion channels (pictured bellow); Leakage channels, voltage-gated channels, ligand-gated channels, and signal-gated channels.
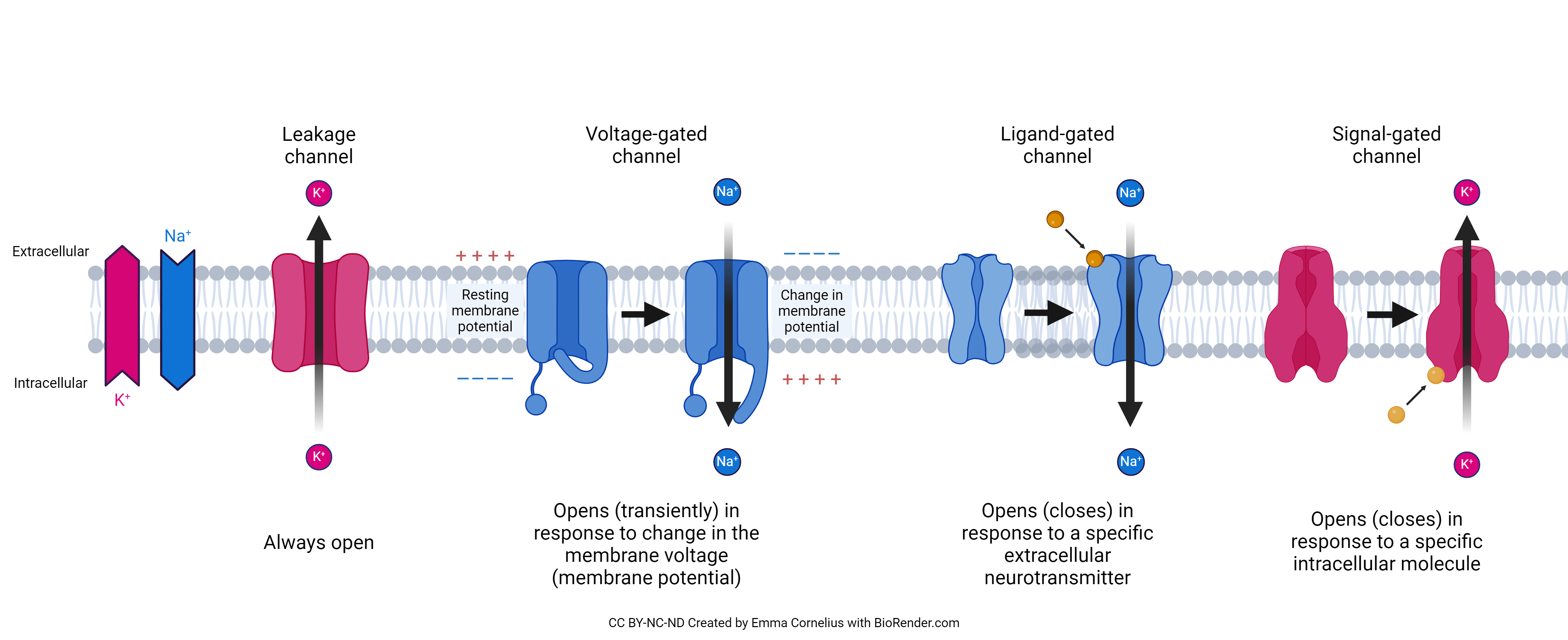
Leakage channels are an ungated channel that allows ion molecules of a certain type to pass through with facilitated diffusion. Potassium (K+) and sodium (Na+) are common for this type of channel. Due to this potassium and sodium are regularly moving across the cell membrane, and play a integral role in the membrane action potention.
Voltage-gated channels will be open or closed depending on the cell membrane’s relative charge (membrane potential). Essentially, they will respond to a change in the membrane potential by altering their shape which either opens, brings to resting close, or inactivates the channel depending on the purpose of the channel.
Ligand-gated channels will open or close depending on the presence of an extracellular ligand. In the case of a neuron, this will be a neurotransmitter.
Signal-gated channels will open or close in response to an intracellular molecule. This is similar to ligand gated channels, with the key difference being what side of the cellular membrane the signaling molecule comes from.
Pumps
Pumps are the type of transporter molecule that solely uses active transport to push molecules against their concentration gradient. The most common pump within neurons is the sodium-potassium pump. This is a primary active transporter which pushes three sodium ions out of the cell along with two potassium ions into the cell (a uniport transporter). There are many more pumps within the membrane, but the sodium-potassium pump plays a vital role in a neurons membrane potential. There are also many pumps which work on other molecules, such as glucose pumps. Proton pumps (within the electron transport chain) are incredibly important for energy metabolism and use within cells.
Other Membrane Proteins
The neuronal cell also has many other signaling proteins that do not transport molecules along the membrane. Cell adhesion molecules are a large group of cell signaling molecules that primarily work to signal between cells. There are three major categories of these molecules, which will be discussed bellow.
The Immunoglobulin Superfamily of cell adhesion molecules are a class which look very similar to immunoglobulins and work to signal the cytoplasm. An example of this is the NCAM and FGF receptor proteins, which both signal for the CREBS cycle through different methods (shown bellow). Neurons will have immune signaling proteins of this superfamily, which tell our immune cells that they are a part of our body, like most other cells in the body. These types of proteins can be peripheral or integral and their exact composition will vary a lot between different people.
[add image or text comparing NCAM and FGF receptor protein pathways]
Cadherins, or Calcium-dependent adherent proteins, form adherent junctions in cells. [Add a bit more about cadherins in neurons]
The neuronal cell will also use Integrin proteins for signaling. These are an integral protein which takes a bent form when inactive, and may be bound on either the inside or outside to cause a conformational change and result in signaling. When bound on the outside a signal is sent into the cell, which will often result in signaling such as cell survival/proliferation and gene expression. When bound to the inside of the cell a signal is sent out of the cell, which will often result in signaling such as cell adhesion and migration.
Media Attributions
- Phospholipid bilayer diagram © Don Fawcett is licensed under a All Rights Reserved license
- Simple Diffusion © Cierra Memphis Barnett is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
