Declarative and Non-Declarative Memory
Dentyn Sacre and Jim Hutchins
Chapter under construction. This is the first draft. If you have questions, or want to help in the writing or editing process, please contact hutchins.jim@gmail.com.
What is declarative and nondeclarative memory?
The importance of memory as it relates to our normal functioning each and every day can hardly be emphasized enough, and it is easy to underestimate just how much processing the brain undergoes each time we are challenged to remember the answer to a question on a test, what happened during dinner last night, or even how to do simple tasks such as drive a car. Despite how fundamental the process of learning and memory retrieval are, scientists have struggled for centuries to figure out what exactly occurs in the brain when we form memories. After careful observation and experimentation, they have divided memory into distinct categories based on their purpose and the respective systems in the brain which are responsible for their encoding and storage.
What scientists have arrived at are two main categories: declarative and nondeclarative memory. Declarative memory consists of facts (semantic memory) and events (episodic memory), which is also called explicit memory, since they are able to be consciously recalled. For example, remembering that Santiago Ramon y Cajal was from Spain or the chicken for dinner last night was overcooked are declarative memories. Nondeclarative memory consists of skill and habits (procedural memory) along with unconscious associations. This type of memory, also called implicit memory, does not require conscious deliberation, such as riding a bike or responding with fear at the sight of a large, menacing dog.
Key Terms Used in Relation to Memory
Encoding – Induction of information into memory, involving learning and relating information to previous knowledge
Consolidation – The process which transfers short-term memory into long-term storage
Storage – Maintenance of a memory over time
Retrieval – Accessing stored information when needed
Declarative memory
Declarative memory is further divided into semantic memory, consisting of facts and abstractions, and episodic memory, consisting of events and contextual information. These memories can be short-term, lasting only minutes or hours and easily disrupted by other information, or long-term, which are more stable and can last anywhere from days to years after being stored. The conversion of short-term memories into long-term memories occurs through a process known as memory consolidation, involving parts of the brain we’ll discuss later. Because declarative memory is able to be consciously recalled, the terms ‘declarative’ and ‘explicit’ memory can be used interchangeably.
Nondeclarative memory
The further division of nondeclarative memory is a bit more complicated. Primarily, it consists of procedural memory for skills and habits, but also includes unconscious associations and emotional responses, often from classical and instrumental conditioning, and priming, a phenomenon where exposure to one stimulus unconsciously or unintentionally influences one’s response to another stimulus. This form of unconscious memory, synonymous with ‘implicit’ memory, seems to predominate in infants. While explicit memory performance increases early in life and declines in old age, implicit memory performance appears more stable throughout the human lifespan.
The study of memory and evidence for the distinction between declarative and nondeclarative memory
Major events in the history of the study of memory
- Socrates – claimed that all learning was simply a remembering of innate knowledge acquired before birth but forgotten, naming the process of recollection “anamnesis.”
- Hermann Ebbinghaus – pioneered the psychological study of memory, removing it from its mysterious position in philosophical epistemology and placing it under scientific scrutiny.
- Georg Müller and Alfons Pilzecker – described how memory consolidates with time. Early memories are susceptible to being altered, while older memories are more resistant to interference.
- William James – distinguished between long-term and short-term memory
- Sergei Korsakoff – first description of a memory disorder, coming to be known as Korsakoff’s syndrome.
- Cognitive Revolution – emerged in the 1960s as a response to the constraints of behaviorism. Psychologists such as Frederic C Bartlett showed how fragile and malleable memory was. Rarely is memory an exact reflection of past events, but rather it is often an imaginative reconstruction painted by the emotional circumstances that are present during its recollection. The cognitive revolution in the 1960s emphasized one part of human behavior that had been overlooked by the earlier behaviorists: that behavior is a product of internal mechanisms and representations that are hard to study from the outside. By merging with biology, psychology and neuroscience developed ways to make inferences about these internal representations.
- Biological Revolution – with advances in imaging technology and the understanding of molecular and cellular biology, scientists developed the ability to map cognitive functions onto brain areas. This had profound implications for the understanding of memory and the location of memory in the brain.
The engram

Initial neuroscience memory researchers attempted to find the location of memories in the brain, what they called an ‘engram’ or ‘memory trace.’ Karl Lashley, an American psychologist who did research on the brain and learning, investigated what happened to the memory of rats who had various parts of the cerebral cortex removed. His findings indicated that deficits in memory were correlated with size of the region removed, rather than location. The rats displayed what is known as retrograde amnesia, the type of amnesia that involves the loss of previous memories. Later research found methodological errors in Lashley’s studies. Since he tested memory using maze tasks, the removal of parts of the cortex associated with motor or sensory processing interferes with maze task performance. Additionally, he did not investigate what happened when subcortical areas were removed. Still, subsequent research shows memory is widely distributed, although not equally distributed.
Research by researchers such as William Scoville, Brenda Milner, and Wilder Penfield showed that the temporal lobe has important implications for memory. Penfield used electrical stimulation in studies to selectively stimulate different parts of the brain. When stimulating the temporal lobe, patients were able to recall long forgotten memories with vivid sensory detail[1]. William Scoville and Brenda Milner worked with an amnesia patient named Henry Molaison, commonly referred to as just H.M., who underwent a bilateral medial temporal lobectomy, removing much of his hippocampi, parahippocampal cortices, rhinal cortices, and amygdalae to stop frequent seizures he had developed as a result of a childhood bicycle accident.

Henry, beyond his contribution to the understanding of the importance of temporal lobe structures for memory, also provided evidence for the division of memory into declarative and nondeclarative categories. For the remainder of his life, though able to remember much of his past prior to the procedure, he was only able to remember new memories as long as his full attention was not diverted, but quickly forgot whatever he had learned once being distracted, indicating an inability to commit short-term memories into long-term storage. Henry was, however, able to develop new nondeclarative memories. For example, he was taught a mirror-drawing skill he had been presented with after the operation, although he claimed each time that he had never done it before.
While medial temporal lobe lesions seem to affect the process of consolidation, the conversion of short-term declarative memories into long lasting constructs, they seem to leave the ability to learn new skills and habits intact. Henry displayed mainly anterograde amnesia, the type which affects the ability to form new memories, though he did experience some deficits in memories of past events of his life.
The creation of declarative memory

Building on the research from Karl Lashley on mice and maze tasks, Donald Hebb laid the foundations for the modern conception of cell assembly and the engram as something widely distributed across the brain, rather than localized to a single region. When recalling past events,
such as a first date with a romantic partner, we combine the various sensations present in that context to create a more complete mental image of the scenario–various structures involving sensation, perception, and emotion work together to form the memory of what outfit our partner was wearing, the scent of the restaurant we may have stopped to get dinner, and the feelings we had toward the experience. This wide reaching memory is not a result of the creation of new neurons, but rather arises from the creation of new connections between neurons from each of the brain regions involved.
Medial temporal lobe and the neocortex
The medial temporal lobe is a complex area of the brain that consists of structures pertinent to primarily the encoding and consolidation aspects of declarative memory, such as the hippocampus, rhinal cortex, and parahippocampal gyrus. The rhinal cortex, adjacent to the rhinal fissure, can be further broken down into two salient structures: the entorhinal and perirhinal cortices.
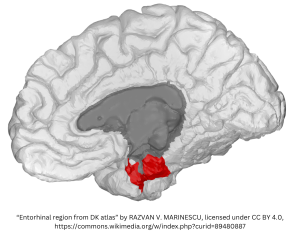
The hippocampus and rhinal cortex are critical for the consolidation of information provided by the association cortex, the part of the cerebral cortex that performs complex cognitive tasks unrelated to sensory or motor processing. After being passed through these structures in the medial temporal lobe, memories then are stored in the neocortex, the outer part of the cerebrum which acts more as a repository for information about the past than a central location for consolidation. Think of the hippocampus and rhinal cortex acting as librarians for the brain, organizing shipments of new books, magazines, and manuscripts into the larger library, the neocortex.
Lesions to the structures of the medial temporal lobe, such as in the case of Henry Molaison, produce anterograde amnesia as the ability to encode and consolidate memory are compromised. Conversely, lesions to the neocortex, such as in Lashley’s rats, produce retrograde amnesia.
Additionally, the hippocampus seems to be responsible for spatial memory and the creation of “cognitive maps,” possibly due to the existence of place cells, special cells in the hippocampus that have been identified in mice and activate only when the organism is in a particular location. Though they have not yet been identified in humans, studies on humans have shown activation in the hippocampus during navigation tasks. Working together with the entorhinal cortex, which provides contextual information, we’re able to form information relevant to “what happened where.” Highlighting the importance of the hippocampus for navigation ability, researchers[2] conducted structural MRI scans on a sample of taxi drivers in London, revealing that they had increased hippocampal volume when compared to a sample of individuals who did not drive taxis. Note that the hippocampus is one of the few brain regions known to continue generating new neurons throughout adulthood.
Consolidation and the engram
Consolidation follows two main types: synaptic consolidation, and systems consolidation. In synaptic consolidation, synapses are remodeled and connections are strengthened as a consequence of activation, producing enduring changes and stabilizing the memory trace. This form occurs rapidly after learning, within minutes or hours, and occurs primarily in the hippocampus. The second type, systems consolidation, is a much slower process, taking up to days or years in the effort to move memories from the locations of consolidation in the medial temporal cortex to distributed areas in the neocortex.
Two theories of consolidation have emerged to answer questions related to the role of the hippocampus. Is the hippocampus necessary for long-term memories? The standard model of consolidation says that the hippocampus is not necessary for long-term retrieval. Under this model, information is sent from the association cortex to areas in the medial temporal lobe where they undergo systems consolidation before being completely relinquished to the neocortex. However, this model does not account for the partial retrograde amnesia that HM displayed. Accordingly, the multiple trace model of consolidation developed and implicates the hippocampus in long-term retrieval. This model states that the neocortex never attains full control over the memory trace or engram, leaving some control in the hands of the hippocampus.
Not only does implicating the hippocampus long-term for memories explain the limited retrograde episodic memory loss that H.M. experienced, but it also has explanatory power for the phenomenon known as reconsolidation. Even after events or experiences are seemingly solidified into our sense of the past, the act of retrieval causes memories to become temporary malleability, and can lead to changes in the memory itself depending on the context we remember it in. This effect may be responsible for memory inaccuracies, such as in eyewitness testimony misidentification cases, where the memory of eyewitnesses is liable to change and conform to information introduced during police questioning due to the enhanced suggestibility within this window provided by reconsolidation. While this temporary malleability can cause changes in memories we perceive as precarious, such as in the case of false memory development or suggestion, the same mechanism of memory instability seems to be the vessel for therapeutic treatment of traumatic disorders or PTSD. In the treatment of PTSD, turning over past memories in a therapeutic setting allows for a reappraisal of the emotional evaluation of past trauma. Another structure, the amygdala, is responsible for the attribution of emotional responses to memory, something that will be explored later.
The creation of nondeclarative memory
The striatum
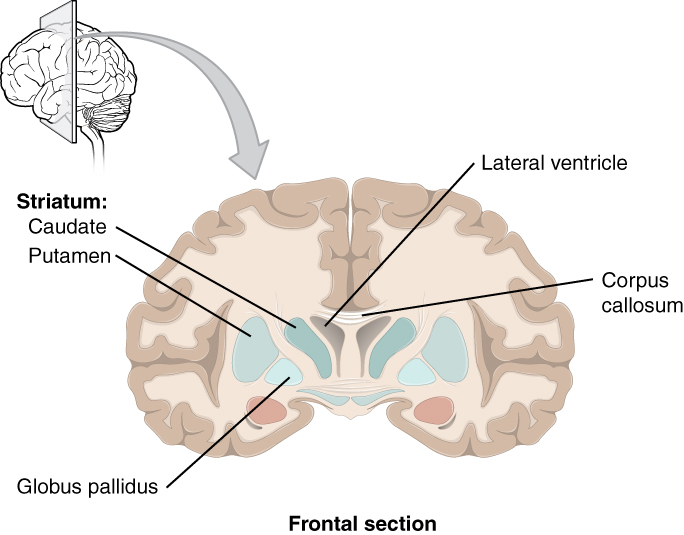
The striatum, a subcortical brain structure consisting of interconnected nuclei and comprising part of the basal ganglia, sits at a location in the motor loop, making it a key player in procedural memory and habit acquisition. It receives input from the frontal and parietal cortices and sends output to thalamic nuclei as well as other cortical areas involved in movement. Its general function includes mediating reward, reinforcement, and stimulus-response learning.
Its role in nondeclarative memory can be understood by comparing amnesia patients like H.M. with Huntington’s patients. In Henry’s case, he was able to learn new procedural tasks despite the devastation he suffered to declarative memory systems, since his striatum remained intact. Huntington’s patients, however, often experience degradation in the striatum as a result of the neurodegenerative nature of the disease. As a result, their declarative memory is left intact, but they struggle in learning tasks where a stimulus is paired with a motor response, independent of motor deficits that plague Huntington’s patients.
The striatum seems to be involved in more than just association of stimuli with motor output. It also deals with associative learning in general. In a study comparing the performance of Parkinson’s patients with amnesic patients on an associative learning task[3], they provided evidence not only for the implication of the striatum in nondeclarative memory, but also supported the distinction between declarative and nondeclarative memory. Parkinson’s patients, who have degeneration of inputs to the striatum from the substantia nigra, showed difficulties in a probabilistic association task, while retaining normal performance on the declarative memory assessment. Conversely, the amnesic patients had poor performance on the declarative task while their performance on the nondeclarative, association task was unaffected.
The connections between the striatum and other brain areas seem to be pertinent to implicit learning as well. The cerebellum, a major motor control center of the brain, is essential for fine motor movement and seems to be a storage location for memory traces related to classical conditioning[4]. The neocortex seems to be involved in the accrual of implicit rules as they relate to categorization or language, making it an essential structure for the application of implicit knowledge in tasks where conscious recollection is not necessary.
Consolidation
Unlike declarative memory, the consolidation of learnt skills such as riding a bike or typing do not require the hippocampus to be stored long-term. Instead, consolidation occurs through the strengthening of connection in brain areas like the striatum, cerebellum, neocortex, and even the amygdala, which is the structure responsible for emotional conditioning.
The amygdala’s role in declarative and nondeclarative memory
The amygdala, often referred to as the emotional processor of the brain, is consequential for both declarative and nondeclarative memory. Its purpose is to associate emotions with memories, making it both a major player in giving emotional context to episodic memories, and the creation of emotional responses to conditioned stimuli, two processes that are closely related and seem to blur the distinction between declarative and nondeclarative memory.
Working together, the amygdala and hippocampus create implicit and explicit aspects of emotional conditioning. The former seems to handle implicit, physiological reactions, such as signaling other regions involved in increased heart rate or fear related hormone release, while the latter allows for explicit recollection of fearful stimuli. In patients with brain damage to the amygdala while the hippocampus is intact, they are able to explicitly recall fearful events or associations, though they lack the physiological indications of conditioned fear. On the other hand, patients with sustained amygdalic damage and spared hippocampi are not able to consciously express fear related to conditioning, but still retain physiological markers of conditioned fear and express aversion to fearful stimuli[5].
Though crucial for implicit fear conditioning, the amygdala’s importance for declarative memory, specifically episodic memory, should not be underscored. The storage of past events is modulated with amygdala-dependent arousal, resulting in the retrieval of long-term memories being influenced by not only the emotion experienced during learning but also the emotion experienced during retrieval. The ease of retrieval of a memory is enhanced by amygdalic involvement, explaining why you might find it easier to remember emotionally charged events, such as a romantic breakup, than emotionally neutral events, like driving to the store. Additionally, if retrieval of a memory occurs in an emotional state similar to the one present during learning, the emotional state may act as a cue that increases the likelihood of retrieval of specific details. Vivid, long lasting memories about emotionally charged events are called flashbulb memories.
Cellular basis of memory
Declarative memory
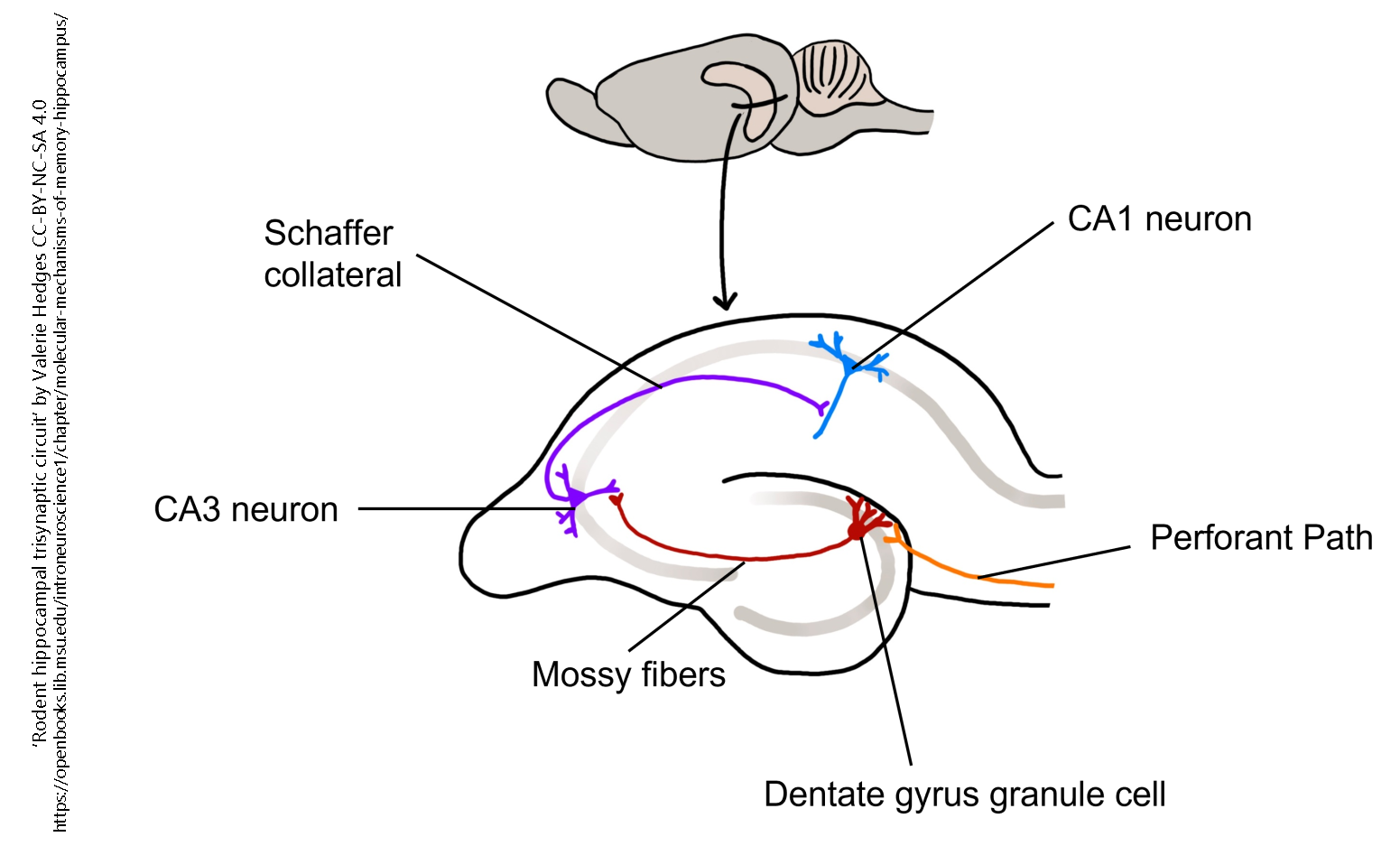
The cellular basis of declarative memory is primarily associated with long-term potentiation and depression, specifically in strengthening connections in the hippocampus and the Schaffer collateral pathway, as well as glutamatergically mediated projections from the hippocampus to other regions of the brain involved in declarative memory.
Schaffer collateral pathway: excitatory glutamatergic pathway in the hippocampus
Long-term potentiation consists of two main processes: early long-term potentiation and late long-term potentiation. Early LTP, completed in only a few hours, does not require protein synthesis. It is limited to changes in the synapse that occur when proteins are activated, resulting in higher sensitivity of AMPA receptors. Additional AMPA receptors may also be recruited to the synapse.
Late LTP, taking days or longer, does involve protein synthesis from the triggering of gene transcription for proteins like BDNF (brain-derived neurotrophic factor) and ARC (activity-regulated cytoskeleton protein). This occurs when a calcium influx from the opening of NMDA receptors activates kinases such as PKA and CaMKII, driving transcription factors like CREB (cAMP response element-binding protein). This results in strengthening of the synapse and structural remodeling that increase receptor density and promote dendritic spine growth. For more information on early- and late-LTP, visit the Introduction to Neuroscience textbook.
Nondeclarative memory

The cellular basis of nondeclarative memory seems to conserve many of the mechanisms of signaling and protein synthesis as seen in declarative memory acquisition, though research is less developed in regards to implicit learning and glutamate seems to play less of a central role. Much of our knowledge of implicit memory originates from studies on invertebrate organisms, since many invertebrate species seem to display exclusively implicit memory. In research done on Aplysia (sea slugs), serotonin seems to be the primary agent in kickstarting protein synthesis, resulting in synaptic strengthening and restructuring[6]. Dopamine pathways involving the striatum also seem to be responsible for habit formation, and it is also suggested that dopamine and acetylcholine independently modulate LTP in learning. Many of these processes occurs at regions involved in nondeclarative memory, such as the striatum, cerebellum, and neocortex
Media Attributions
- Entorhinal cortex is licensed under a CC BY (Attribution) license
- Constudoverbrain
- Aplysia californica
- Kumar, R., & Yeragani, V. K. (2011) Penfield – A great explorer of psyche-soma-neuroscience. Indian Journal of Psychiatry, 53(3). 276-278. https://doi.org/10.4103/0019-5545.86826 ↵
- Maguire E. A., Guardian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R. S. J., & Frith, C. D. (2000). Navigation-related structural changes in the hippocampi of taxi drivers. PNAS, 97(8), 4398-4403. https://doi.org/10.1073/pnas.070039597 ↵
- Knowlton, B. J., Mangels, J. A., & Squire, L. R. A neostriatal habit learning system in humans. Science, 273(5280), 1399-1402. https://doi.org/10.1126/science.273.5280.1399 ↵
- Squire, L. R. (2009). Memory and brain systems: 1969-2009. Journal of Neuroscience, 29(41), 12711-12716. https://doi.org/10.1523/JNEUROSCI.3575-09.2009 ↵
- Phelps, E. A., & LeDoux, J. E. (2005). Contributions of the amygdala to emotion processing: From animal models to human behavior. Neuron, 48(2), 175-187. https://doi.org/10.1016/j.neuron.2005.09.025 ↵
- Kandel, E. R. (2012). The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Molecular Brain, 5(14). https://doi.org/10.1186/1756-6606-5-14; Barco, A., Bailey, C, H., & Kandel, E. R. (2006). Common molecular mechanisms in explicit and implicit memory. Journal of Neurochemistry, 97(6), 1520-1533. https://doi.org/10.1111/j.1471-4159.2006.03870.x ↵
