11.4 Speciation
Learning Objectives
By the end of this section, you will be able to:
- Describe the definition of species and how species are identified as different
- Explain allopatric and sympatric speciation
- Describe adaptive radiation
The biological definition of species, which works for sexually reproducing organisms, is a group of actually or potentially interbreeding individuals. According to this definition, one species is distinguished from another by the possibility of matings between individuals from each species to produce fertile offspring. There are exceptions to this rule. Many species are similar enough that hybrid offspring are possible and may often occur in nature, but for the majority of species this rule generally holds. In fact, the presence of hybrids between similar species suggests that they may have descended from a single interbreeding species and that the speciation process may not yet be completed.
Given the extraordinary diversity of life on the planet there must be mechanisms for speciation: the formation of two species from one original species. Darwin envisioned this process as a branching event and diagrammed the process in the only illustration found in On the Origin of Species (Figure 11.14a). For speciation to occur, two new populations must be formed from one original population, and they must evolve in such a way that it becomes impossible for individuals from the two new populations to interbreed. Biologists have proposed mechanisms by which this could occur that fall into two broad categories. Allopatric speciation, meaning speciation in “other homelands,” involves a geographic separation of populations from a parent species and subsequent evolution. Sympatric speciation, meaning speciation in the “same homeland,” involves speciation occurring within a parent species while remaining in one location.
Biologists think of speciation events as the splitting of one ancestral species into two descendant species. There is no reason why there might not be more than two species formed at one time except that it is less likely and such multiple events can also be conceptualized as single splits occurring close in time.

Speciation through Geographic Separation
A geographically continuous population has a gene pool that is relatively homogeneous. Gene flow, the movement of alleles across the range of the species, is relatively free because individuals can move and then mate with individuals in their new location. Thus, the frequency of an allele at one end of a distribution will be similar to the frequency of the allele at the other end. When populations become geographically discontinuous that free-flow of alleles is prevented. When that separation lasts for a period of time, the two populations are able to evolve along different trajectories. Thus, their allele frequencies at numerous genetic loci gradually become more and more different as new alleles independently arise by mutation in each population. Typically, environmental conditions, such as climate, resources, predators, and competitors, for the two populations will differ causing natural selection to favor divergent adaptations in each group. Different histories of genetic drift, enhanced because the populations are smaller than the parent population, will also lead to divergence.
Given enough time, the genetic and phenotypic divergence between populations will likely affect characters that influence reproduction enough that were individuals of the two populations brought together, mating would be less likely, or if a mating occurred, offspring would be non-viable or infertile. Many types of diverging characters may affect the reproductive isolation (inability to interbreed) of the two populations. These mechanisms of reproductive isolation can be divided into prezygotic mechanisms (those that operate before fertilization) and postzygotic mechanisms (those that operate after fertilization). Prezygotic mechanisms include traits that allow the individuals to find each other, such as the timing of mating, sensitivity to pheromones, or choice of mating sites. If individuals are able to encounter each other, character divergence may prevent courtship rituals from leading to a mating either because female preferences have changed or male behaviors have changed. Physiological changes may interfere with successful fertilization if mating is able to occur. Postzygotic mechanisms include genetic incompatibilities that prevent proper development of the offspring, or if the offspring live, they may be unable to produce viable gametes themselves as in the example of the mule, the infertile offspring of a female horse and a male donkey.
If the two isolated populations are brought back together and the hybrid offspring that formed from matings between individuals of the two populations have lower survivorship or reduced fertility, then selection will favor individuals that are able to discriminate between potential mates of their own population and the other population. This selection will enhance the reproductive isolation.
Isolation of populations leading to allopatric speciation can occur in a variety of ways: from a river forming a new branch, erosion forming a new valley, or a group of organisms traveling to a new location without the ability to return, such as seeds floating over the ocean to an island. The nature of the geographic separation necessary to isolate populations depends entirely on the biology of the organism and its potential for dispersal. If two flying insect populations took up residence in separate nearby valleys, chances are that individuals from each population would fly back and forth, continuing gene flow. However, if two rodent populations became divided by the formation of a new lake, continued gene flow would be unlikely; therefore, speciation would be more likely.
Biologists group allopatric processes into two categories. If a few members of a species move to a new geographical area, this is called dispersal. If a natural situation arises to physically divide organisms, this is called vicariance.
Scientists have documented numerous cases of allopatric speciation taking place. For example, along the west coast of the United States, two separate subspecies of spotted owls exist. The northern spotted owl has genetic and phenotypic differences from its close relative, the Mexican spotted owl, which lives in the south (Figure 11.15). The cause of their initial separation is not clear, but it may have been caused by the glaciers of the ice age dividing an initial population into two.1
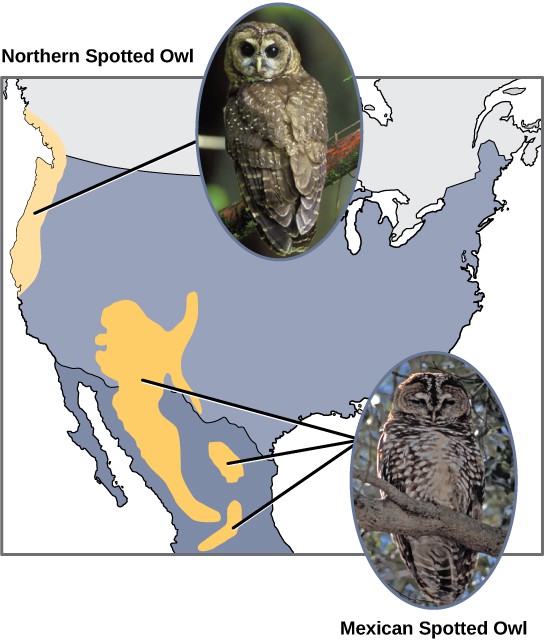
Additionally, scientists have found that the further the distance between two groups that once were the same species, the more likely for speciation to take place. This seems logical because as the distance increases, the various environmental factors would likely have less in common than locations in close proximity. Consider the two owls; in the north, the climate is cooler than in the south; the other types of organisms in each ecosystem differ, as do their behaviors and habits; also, the hunting habits and prey choices of the owls in the south vary from the northern ones. These variances can lead to evolved differences in the owls, and over time speciation will likely occur unless gene flow between the populations is restored.
In some cases, a population of one species disperses throughout an area, and each finds a distinct niche or isolated habitat. Over time, the varied demands of their new lifestyles lead to multiple speciation events originating from a single species, which is called adaptive radiation. From one point of origin, many adaptations evolve causing the species to radiate into several new ones. Island archipelagos like the Hawaiian Islands provide an ideal context for adaptive radiation events because water surrounds each island, which leads to geographical isolation for many organisms (Figure 11.16). The Hawaiian honeycreeper illustrates one example of adaptive radiation. From a single species, called the founder species, numerous species have evolved, including the eight shown in Figure 11.16.
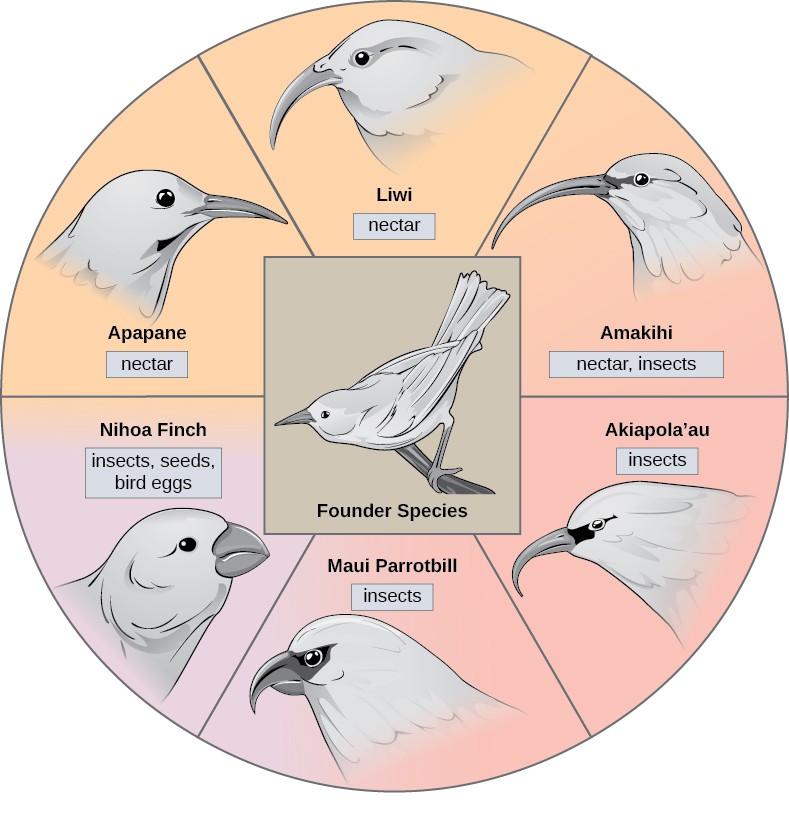
Notice the differences in the species’ beaks in Figure 11.16]. Change in the genetic variation for beaks in response to natural selection based on specific food sources in each new habitat led to evolution of a different beak suited to the specific food source. The fruit and seed-eating birds have thicker, stronger beaks which are suited to break hard nuts. The nectar-eating birds have long beaks to dip into flowers to reach their nectar. The insect-eating birds have beaks like swords, appropriate for stabbing and impaling insects. Darwin’s finches are another well-studied example of adaptive radiation in an archipelago.
Concept in Action

Speciation without Geographic Separation
Can divergence occur if no physical barriers are in place to separate individuals who continue to live and reproduce in the same habitat? A number of mechanisms for sympatric speciation have been proposed and studied.
One form of sympatric speciation can begin with a chromosomal error during meiosis or the formation of a hybrid individual with too many chromosomes. Polyploidy is a condition in which a cell, or organism, has an extra set, or sets, of chromosomes. Scientists have identified two main types of polyploidy that can lead to reproductive isolation of an individual in the polyploid state. In some cases a polyploid individual will have two or more complete sets of chromosomes from its own species in a condition called autopolyploidy (Figure 11.17). The prefix “auto” means self, so the term means multiple chromosomes from one’s own species. Polyploidy results from an error in meiosis in which all of the chromosomes move into one cell instead of separating.
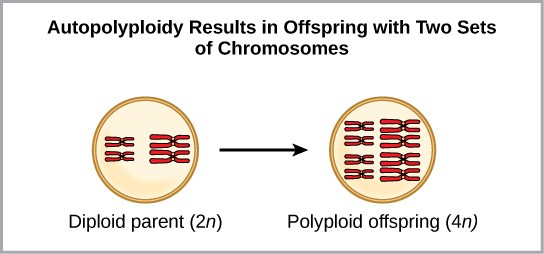
For example, if a plant species with 2n = 6 produces autopolyploid gametes that are also diploid (2n = 6, when they should be n = 3), the gametes now have twice as many chromosomes as they should have. These new gametes will be incompatible with the normal gametes produced by this plant species. But they could either self-pollinate or reproduce with other autopolyploid plants with gametes having the same diploid number. In this way, sympatric speciation can occur quickly by forming offspring with 4n called a tetraploid. These individuals would immediately be able to reproduce only with those of this new kind and not those of the ancestral species. The other form of polyploidy occurs when individuals of two different species reproduce to form a viable offspring called an allopolyploid. The prefix “allo” means “other” (recall from allopatric); therefore, an allopolyploid occurs when gametes from two different species combine. Figure 11.18 illustrates one possible way an allopolyploidy can form. Notice how it takes two generations, or two reproductive acts, before the viable fertile hybrid results.
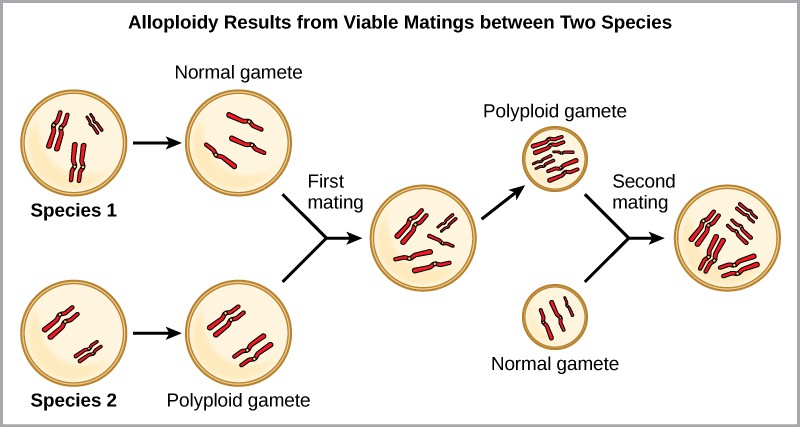
The cultivated forms of wheat, cotton, and tobacco plants are all allopolyploids. Although polyploidy occurs occasionally in animals, most chromosomal abnormalities in animals are lethal; it takes place most commonly in plants. Scientists have discovered more than 1/2 of all plant species studied relate back to a species evolved through polyploidy.
Sympatric speciation may also take place in ways other than polyploidy. For example, imagine a species of fish that lived in a lake. As the population grew, competition for food also grew. Under pressure to find food, suppose that a group of these fish had the genetic flexibility to discover and feed off another resource that was unused by the other fish. What if this new food source was found at a different depth of the lake? Over time, those feeding on the second food source would interact more with each other than the other fish; therefore they would breed together as well. Offspring of these fish would likely behave as their parents and feed and live in the same area, keeping them separate from the original population. If this group of fish continued to remain separate from the first population, eventually sympatric speciation might occur as more genetic differences accumulated between them.
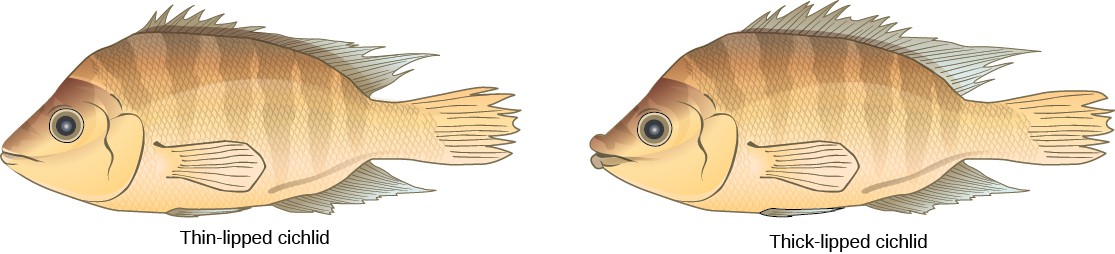
This scenario does play out in nature, as do others that lead to reproductive isolation. One such place is Lake Victoria in Africa, famous for its sympatric speciation of cichlid fish. Researchers have found hundreds of sympatric speciation events in these fish, which have not only happened in great number, but also over a short period of time. Figure 11.19 shows this type of speciation among a cichlid fish population in Nicaragua. In this locale, two types of cichlids live in the same geographic location; however, they have come to have different morphologies that allow them to eat various food sources.
Finally, a well-documented example of ongoing sympatric speciation occurred in the apple maggot fly, Rhagoletis pomonella, which arose as an isolated population sometime after the introduction of the apple into North America. The native population of flies fed on hawthorn species and is host-specific: it only infests hawthorn trees. Importantly, it also uses the trees as a location to meet for mating. It is hypothesized that either through mutation or a behavioral mistake, flies jumped hosts and met and mated in apple trees, subsequently laying their eggs in apple fruit. The offspring matured and kept their preference for the apple trees effectively dividing the original population into two new populations separated by host species, not by geography. The host jump took place in the nineteenth century, but there are now measureable differences between the two populations of fly. It seems likely that host specificity of parasites in general is a common cause of sympatric speciation.
Section Summary
Speciation occurs along two main pathways: geographic separation (allopatric speciation) and through mechanisms that occur within a shared habitat (sympatric speciation). Both pathways force reproductive isolation between populations. Sympatric speciation can occur through errors in meiosis that form gametes with extra chromosomes, called polyploidy. Autopolyploidy occurs within a single species, whereas allopolyploidy occurs because of a mating between closely related species. Once the populations are isolated, evolutionary divergence can take place leading to the evolution of reproductive isolating traits that prevent interbreeding should the two populations come together again. The reduced viability of hybrid offspring after a period of isolation is expected to select for stronger inherent isolating mechanisms.
Glossary
- adaptive radiation: a speciation when one species radiates out to form several other species
- allopatric speciation: a speciation that occurs via a geographic separation
- dispersal: an allopatric speciation that occurs when a few members of a species move to a new geographical area
- speciation: a formation of a new species
- sympatric speciation: a speciation that occurs in the same geographic space
- vicariance: an allopatric speciation that occurs when something in the environment separates organisms of the same species into separate groups
Footnotes
- 1 Courtney, S.P., et al, “Scientific Evaluation of the Status of the Northern Spotted Owl,” Sustainable Ecosystems Institute (2004), Portland, OR.
Media Attributions
- Figure 11.14 © OpenStax is licensed under a CC BY (Attribution) license
- Figure 11.15 © (A) “Northern Spotted Owl” Modification of work by John and Karen Hollingsworth, USFWS; (B) “Mexican Spotted Owl” Modification of work by Bill Radke, USFWS; OpenStax is licensed under a CC BY (Attribution) license
- Figure 11.16 © OpenStax is licensed under a CC BY (Attribution) license
- 11.4QR
- Figure 11.17 © OpenStax is licensed under a CC BY (Attribution) license
- Figure 11.18 © OpenStax is licensed under a CC BY (Attribution) license
- Figure 11.19 © OpenStax is licensed under a CC BY (Attribution) license
