The Adaptive Immune System
Objective 11
Explain the difference between cell-mediated and antibody-mediated immunity. Describe how antigen presentation by MHC Class I molecules activates the adaptive immune system. Describe how antigen presentation by MHC Class II molecules activates another set of responses in the adaptive immune system. Recall how B cells are recruited by T helper cells.
Adaptive immunity can be divided into cell-mediated and antibody-mediated immunity. Both mechanisms are triggered by exposure to specific antigens.
In cell-mediated immunity, T lymphocytes are activated directly against abnormal cells, such as cancer cells, transplanted tissues or infected body cells.
In antibody-mediated immunity, B lymphocytes are recruited and activated by T helper cells to become plasma B cells, which produce and secrete specific antibodies. Antibodies are found in the plasma as well as other fluids throughout the body, which is why an older name for antibody mediated immunity is humoral immunity.
Because T helper cells are the middlemen of adaptive immunity, they play a part in both cell-mediated and antibody-mediated immunity.
Lymphocytes have specificity. Remember that each lymphocyte reacts only to one of the 100,000,000 possible antigens. This means that the cell will spend its life fighting the specific antigen that activated it. For example, if a B cell recognizes a specific protein on the surface of the hepatitis B virus, it will continue to produce antibodies to that one specific protein on that one specific virus. If the cell divides to form more plasma B cells or memory B cells, the daughter cells will have the same genetic makeup of the activated parent B cell (i.e. are clones) and will retain specificity to that same antigen.
We call these identical copies of the same cell a monoclonal population, and the antibodies made by these cells are called monoclonal antibodies. Through this, you are able to have cells and antibodies that help protect you against pathogens you may not have been exposed to for years or even decades.
Antigen Presentation by Infected Cancerous Cells (MHC Class I Markers)
 MHC Class I molecules are found on the surface of all nucleated cells. That’s because any cell with DNA in its nucleus is subject to having that DNA altered by a virus, or by a mutation event which triggers cancer.
MHC Class I molecules are found on the surface of all nucleated cells. That’s because any cell with DNA in its nucleus is subject to having that DNA altered by a virus, or by a mutation event which triggers cancer.
In each of these cases, the change in DNA means that a different set of proteins, or unusual forms of the usual proteins, will be made by the cell. The cell’s proteosomes (Unit 5) are busy shredding all the proteins of the cell, and then small pieces of each protein made by the cell are displayed on the cell surface.
The MHC molecules then act as “billboards” to display the protein fragments.
As long as the protein fragments are the same ones the immune system has always seen, nothing much happens.
But if the protein fragments displayed on the MHC Class I billboard are weird, then the T cytotoxic cell with specificity for that particular kind of weirdness springs into action.
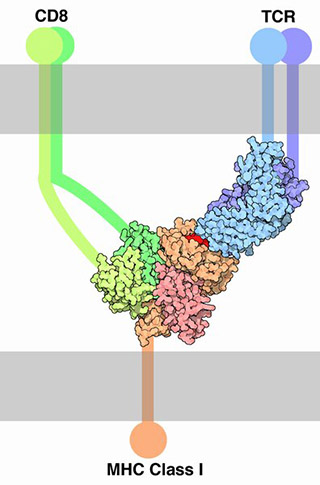
Note the complex of MHC Class I marker, T cell receptor (TCR), and CD8 marker illustrated here. Let’s break down its components:
- an antigen (red);
- a sort-of half-antibody molecule (blue) which is part of the TCR complex, and is specific for the red antigen molecule;
- the T cell marker CD3 we discussed earlier (purple) which is also part of the TCR complex;
- the CD8 marker which is specific to T cytotoxic cells (green).
Since CD8 markers are found only on T cytotoxic (TC) lymphocytes, this complex only binds to T cytotoxic cells. When the complex shown here forms, the T cytotoxic cell gets turned on, and when it’s turned on it releases two chemical factors that destroy the weird-looking cell that originally displayed its weirdness.
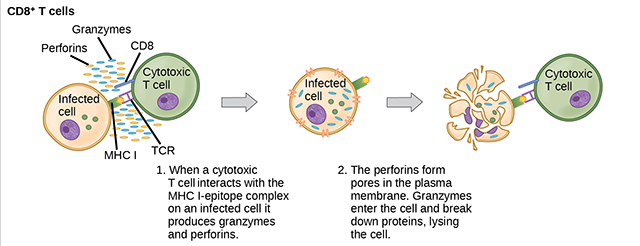
Granzymes are chemical signals that activate an apoptotic program, and the cell dies by digesting itself until the macrophage comes along to clean up the mess left behind.
Perforin, like the name suggests, perforates the weird cell’s membrane. It turns out perforin is almost identical to complement C9, part of the membrane attack complex we saw in Objective 10. That makes sense, because both perforin and the MAC do the same thing: punch holes in the weird cell membrane (or invader cell membrane) until it bleeds out and dies.
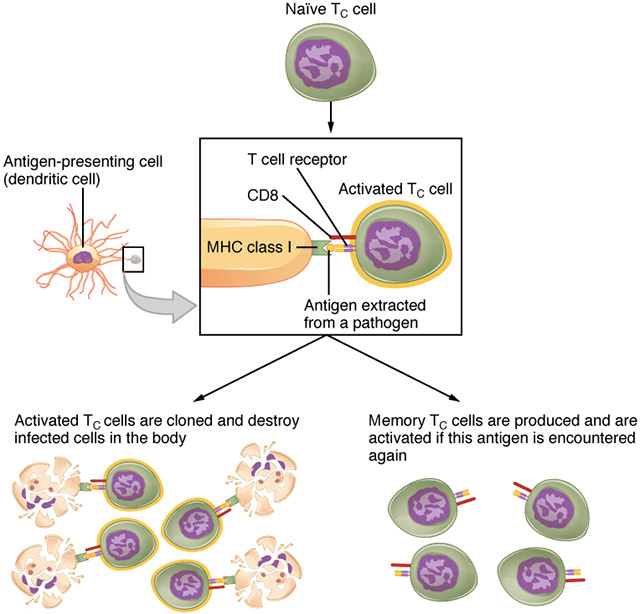
It doesn’t end there. If a dendritic cell picks up the unusual protein fragment that triggered this whole thing, then it displays that weird antigen on its own MHC I billboard. This not only recruits any naïve (previously unactivated) T cytotoxic cells that have this specific antigen-binding site, but also causes the T cytotoxic cells to divide and form memory T cytotoxic cells so there are thousands of these trained killers available next time you encounter this antigen.
Antigen Presentation by Antigen Presenting Cells (MHC Class II Markers)
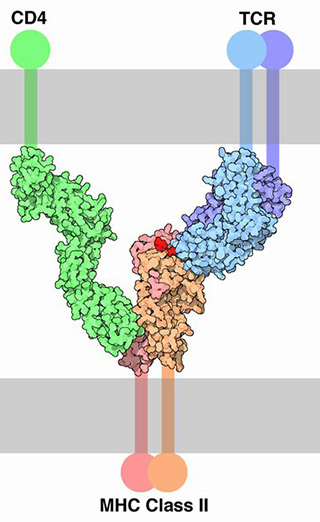 Now, carefully examine the structure of the MHC Class II complex. There’s the antigen (red), held by that half-antibody (blue). The blue protein is paired with CD3 (purple) to form the TCR. The MHC Class II molecule binds to CD4 (green), so the cell it’s binding to, and activating, is now a T helper (TH) cell.
Now, carefully examine the structure of the MHC Class II complex. There’s the antigen (red), held by that half-antibody (blue). The blue protein is paired with CD3 (purple) to form the TCR. The MHC Class II molecule binds to CD4 (green), so the cell it’s binding to, and activating, is now a T helper (TH) cell.
MHC Class II molecules are used by “professional” antigen-presenting cells, the dendritic cells and macrophages, for antigen display.
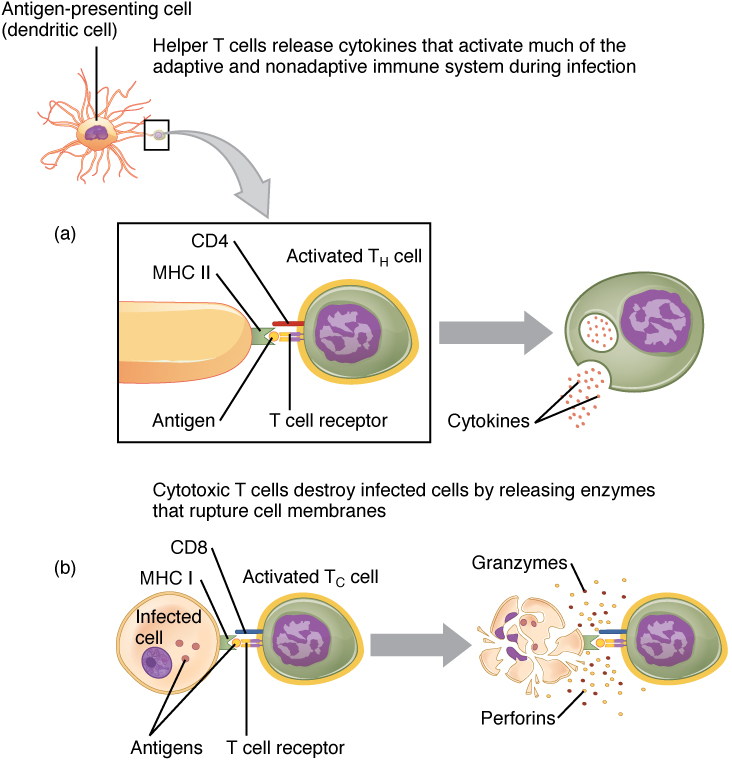
For example, the dendritic cell acts as a sort of war correspondent. It travels to areas where the immune system is locked in a fight with invaders or weird cells, and takes a “photograph” of the war zone by picking up pieces of damaged cells and taking them to the lymph nodes. Once in the lymph node, the dendritic cell plasters those enemy antigens on its MHC Class II billboard and recruits T helper cells to come fight.
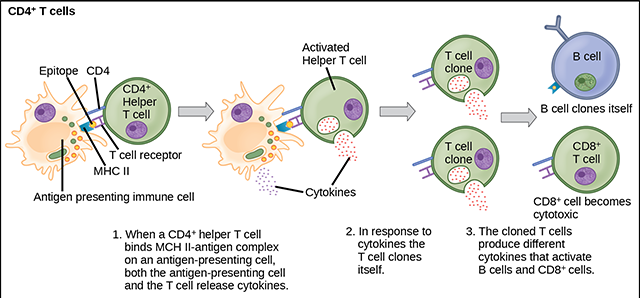
The activated T helper cell does this by releasing cytokine signals, and the cytokines then stimulate a whole variety of immune cells to help in the battle. Notably, the T helper cell makes exact copies of itself (clones) and the clones release cytokines that attract TC and B cells, which then clone themselves. In this way, the immune system creates thousands of cells with different talents which are specialized against the target originally discovered by the dendritic cell or macrophage.
Antigen Presentation by Experienced B Cells (MHC Class II Markers)
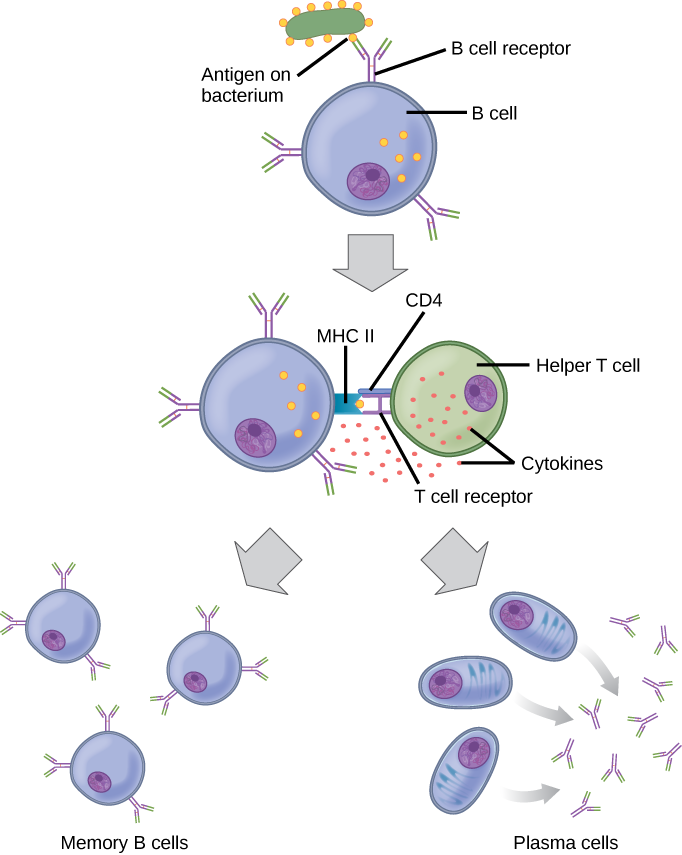
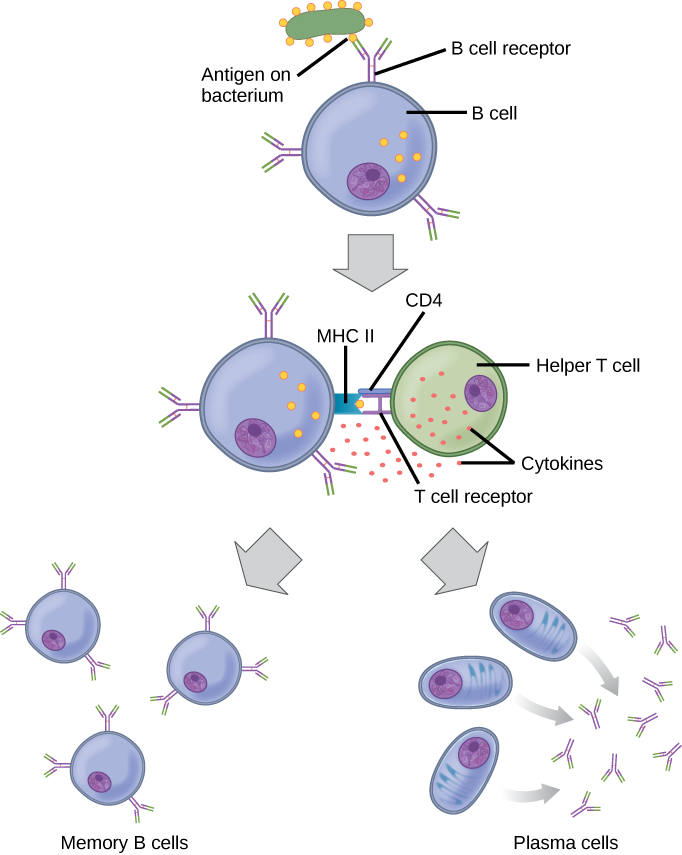
Naïve B cells (i.e. those that have not been exposed to their antigen) don’t present antigen. But an activated B cell can also act as an antigen-presenting cell.
It does this by ramping up the expression of MHC class II molecules which hold onto, and display, the antigen they’re specific for. Remember that the MHC class II molecule also has binding sites for CD4 and the TCR, which together define a T helper cell. The T helper releases cytokines that cause the B cell to divide; some of the clones become plasma cells, and some become memory B cells.
Media Attributions
- U15-075 T Cell Receptor MHC Class I © Goodsell, David adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- U15-076 T Cytotixic Cell © OpenStax adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- U15-077 Clonal Selection and Expression of T Lymphocytes © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license
- U15-078 T Cell Receptor MHC Class II © Goodsell, David adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- U15-079 Pathogen Presentation © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license
- U15-080 CD4 T Cells © OpenStax adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- U15-081 Binding of MHC Class II © Molnar, Charles and Gair, Jane is licensed under a CC BY (Attribution) license

{kind=link}