Sensory Systems | Somatosensory (Touch and Body Position) System
Objective 4
List the elements of the somatosensory (touch) system: receptors, their pathways into the central nervous system, and their pathways within the central nervous system. Understand how touch information is transduced in different receptor types.
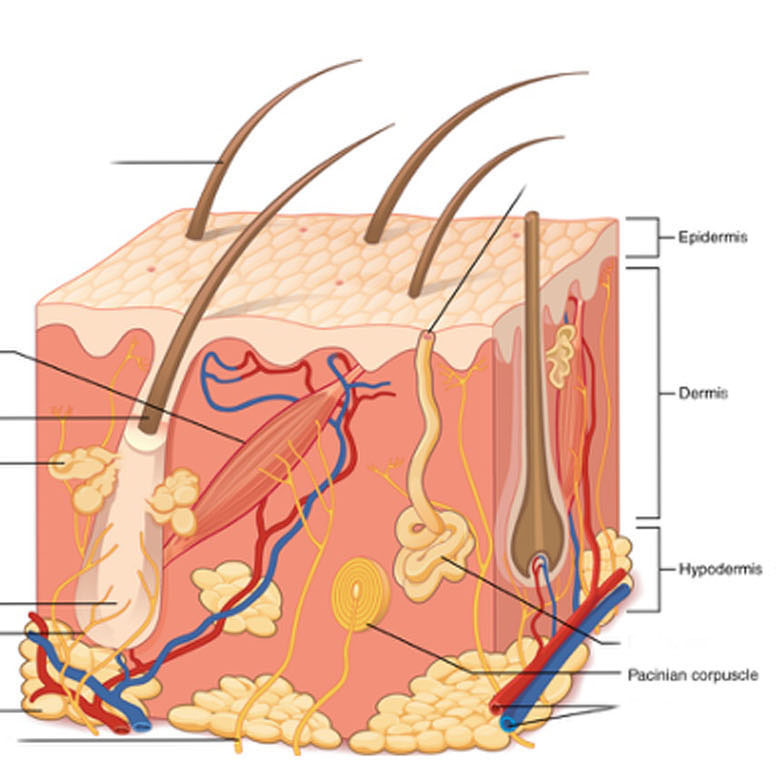
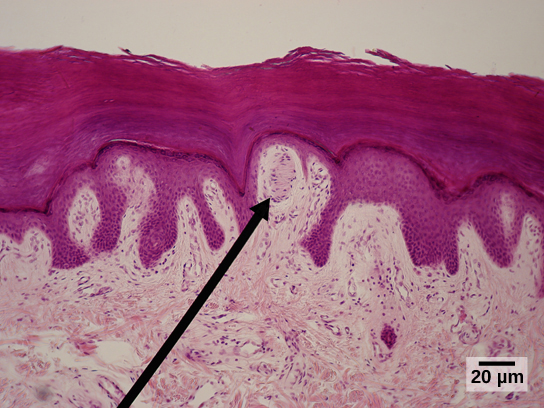
Remember from Unit 8 the layers of the skin: the epidermis, whose basal layer consists of stem cells that give rise to the dying and dead cells (keratinocytes) making up most of the skin thickness; and the dermis, the living connective tissue layer where blood vessels and most receptors are located.
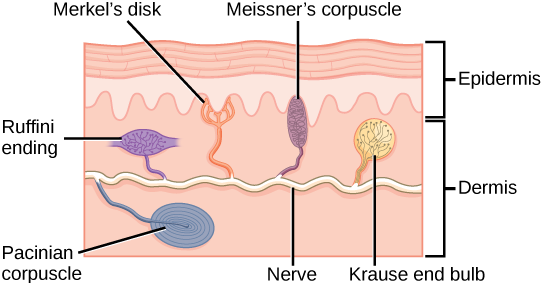
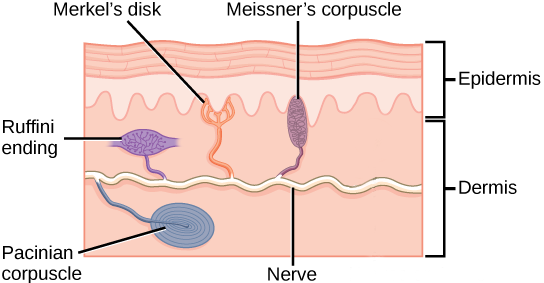 Different authorities may count them differently, but in this course, we will list five different types of receptor. Four of them are shown in the image at right.
Different authorities may count them differently, but in this course, we will list five different types of receptor. Four of them are shown in the image at right.
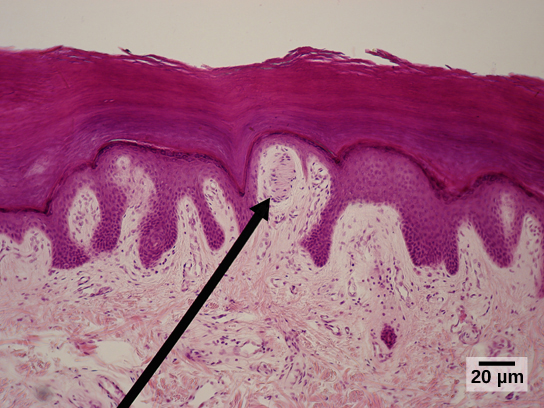
- Merkel disks, whose sensory endings are in the epidermis as we saw in Unit 8. In keeping with their location, these receptors sense light touch, tiny deflections of the skin surface;
 Meissner corpuscles, also carrying a different aspect of the light touch sensation, in the dermis just deep to the basal layer of the epidermis;
Meissner corpuscles, also carrying a different aspect of the light touch sensation, in the dermis just deep to the basal layer of the epidermis;
- Ruffini endings, deeper in the dermis and detecting larger skin deflections (crude touch);
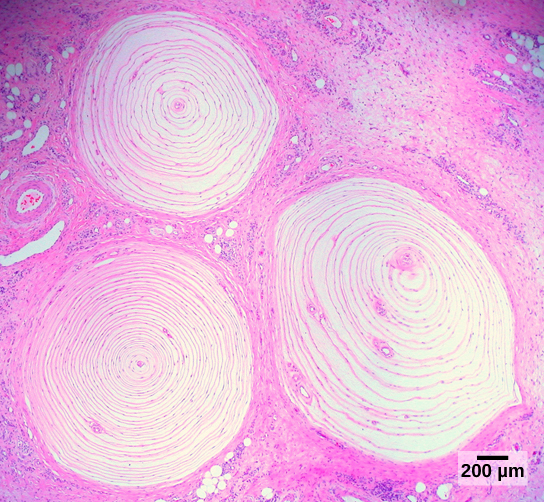
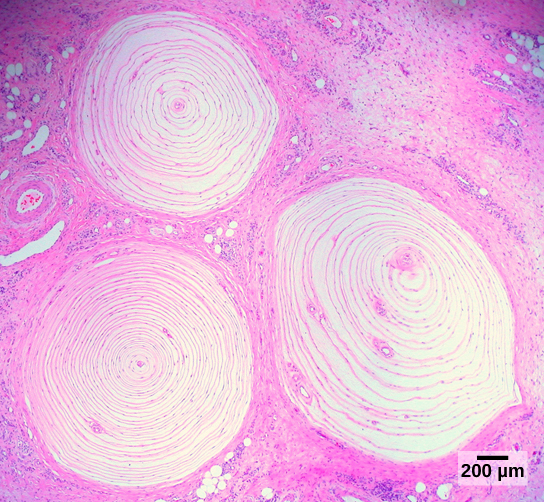 Pacinian corpuscles, also deep in the dermis but able to respond to rapid, repetitive deflections of the skin (vibration);
Pacinian corpuscles, also deep in the dermis but able to respond to rapid, repetitive deflections of the skin (vibration);
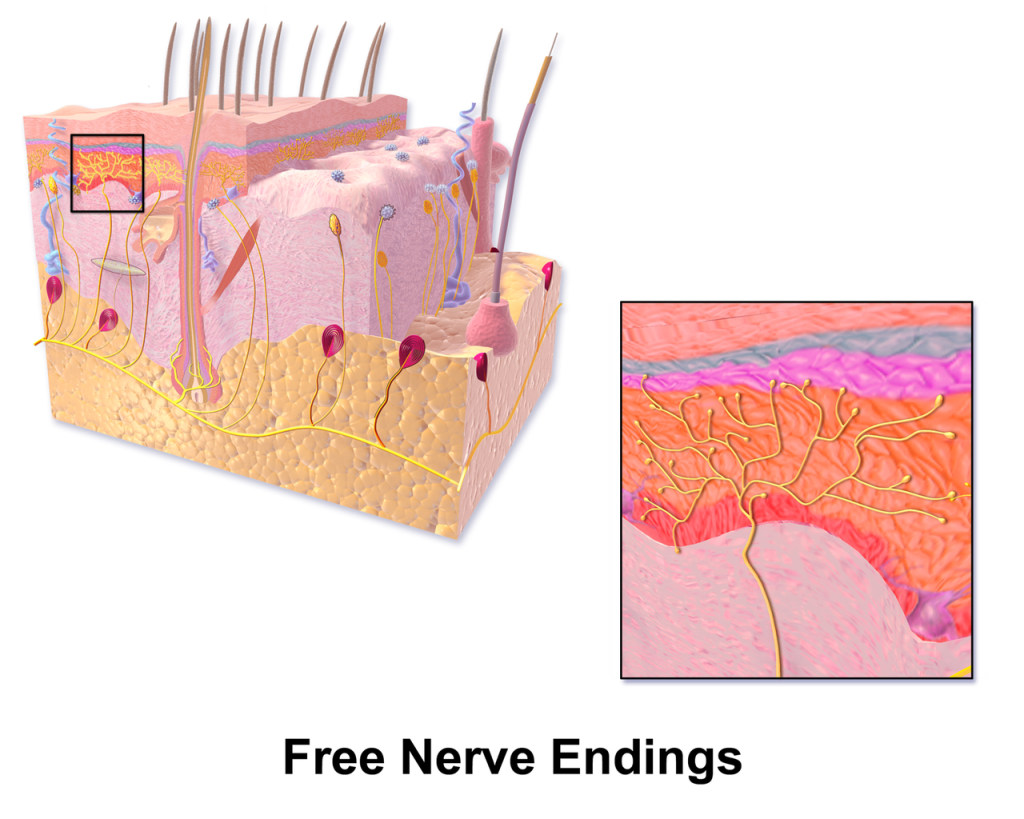 free nerve endings, found at all depths of the dermis (not shown in this figure), some types transducing pain information (nociceptors), some types transducing cold (4°C to 37°C) and some types transducing heat (37°C to 50°C). Beyond these temperature ranges, heat and cold are perceived as pain. There are also free nerve endings responsible for itch, but these will not be discussed in this course.
free nerve endings, found at all depths of the dermis (not shown in this figure), some types transducing pain information (nociceptors), some types transducing cold (4°C to 37°C) and some types transducing heat (37°C to 50°C). Beyond these temperature ranges, heat and cold are perceived as pain. There are also free nerve endings responsible for itch, but these will not be discussed in this course.
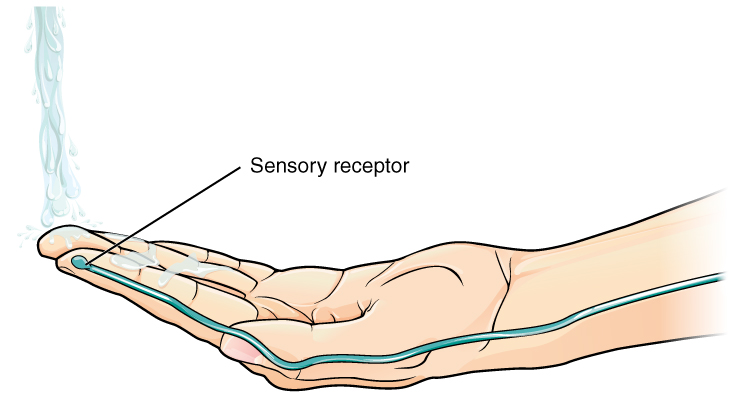 Sensory neurons, by definition, receive information from the environment. Therefore, when the relevant stimulus is received, it causes a change in voltage inside the cell. Neuroscientists call voltage “potential” (because in physics, voltage represents electrical potential energy and current represents electrical kinetic energy). For that reason, the voltage changed triggered by an environmental stimulus is called the generator potential.
Sensory neurons, by definition, receive information from the environment. Therefore, when the relevant stimulus is received, it causes a change in voltage inside the cell. Neuroscientists call voltage “potential” (because in physics, voltage represents electrical potential energy and current represents electrical kinetic energy). For that reason, the voltage changed triggered by an environmental stimulus is called the generator potential.
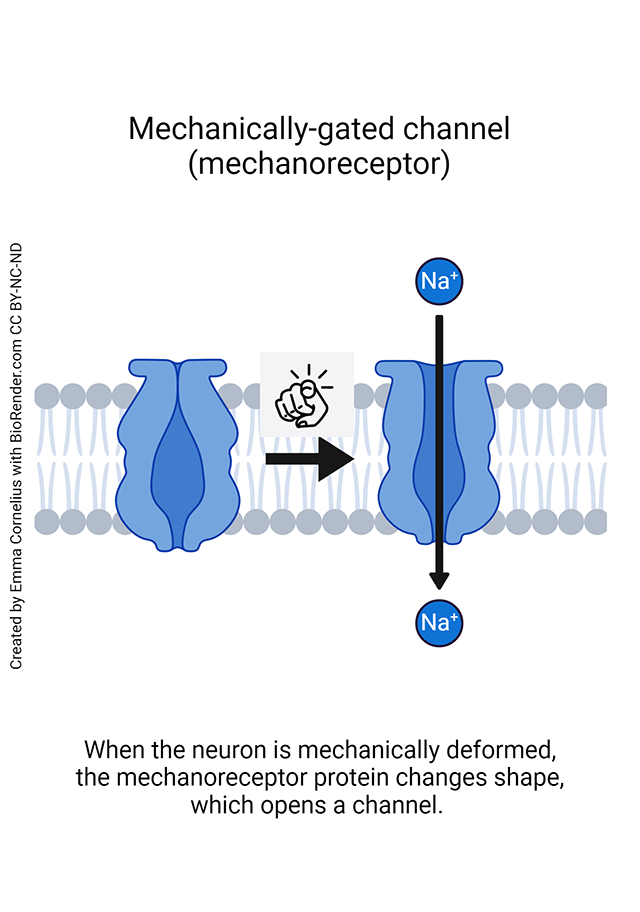 Generator potentials are initiated by mechanoreceptors, proteins that change shape when under pressure and then begin allowing positively-charged sodium ions (Na+) into the nerve cell. This makes the inside of the receptor neuron more positive. This positive voltage deflection, in turn, might generate an action potential (as we saw for muscle fibers in Unit 10), an electrical wave that can travel along nerve axons.
Generator potentials are initiated by mechanoreceptors, proteins that change shape when under pressure and then begin allowing positively-charged sodium ions (Na+) into the nerve cell. This makes the inside of the receptor neuron more positive. This positive voltage deflection, in turn, might generate an action potential (as we saw for muscle fibers in Unit 10), an electrical wave that can travel along nerve axons.
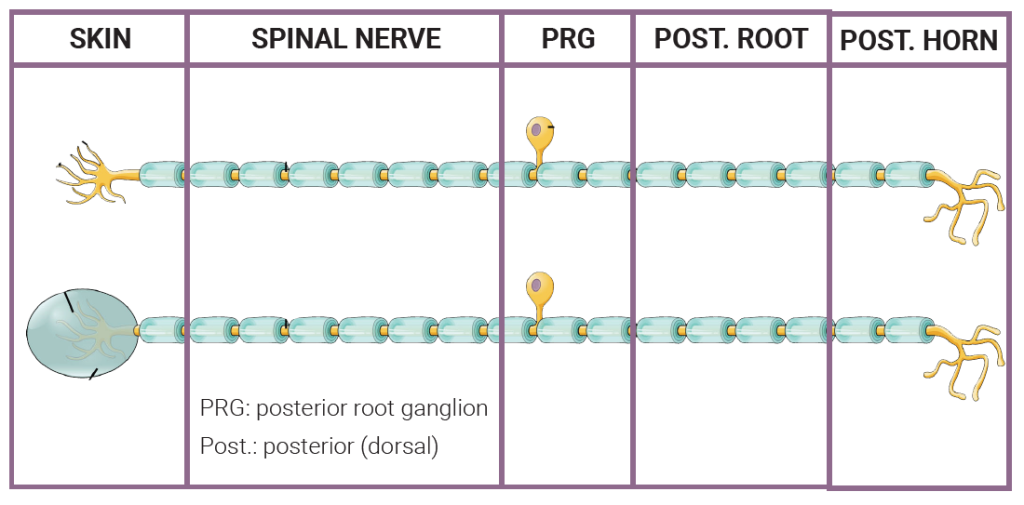
In Unit 11, we described three shapes of neurons. Sensory cells in the skin fit into the category called unipolar neurons. This means they have a dendrite which picks up information, a long axon which starts at the base of the dendritic tree and extends into the CNS, and a cell body shoved off to one side somewhere in between.
The generator potential arises in the dendritic tree. The cell body is in a dorsal root ganglion (called posterior root ganglion in humans). The axon terminals of nociceptors and thermoreceptors are in the dorsal horn of the spinal cord. The axon terminals of mechanoreceptors are in the medulla.
Remember, there are many types of mechanoreceptors, each with a relevant stimulus that’s unique to that cell type. The specializations around the modified dendrite, such as the disk of Merkel discs or the onionskin layers of the Pacinian corpuscle alter the way in which the stimulus is received. (Scientists have experimentally removed these endings and when they do, these receptors all act the same.)
Pain receptor proteins on nociceptors and temperature receptor proteins on thermoreceptors are less understood. Thermoreceptors work because of a change in the shape of temperature-sensitive transmembrane proteins called TRP proteins. TRP proteins also bind to capsaicin (the “hot” in hot peppers) and menthol (the “cool” in mint). When we describe those chemicals as “hot” or “cold”, then, there is a real molecular basis for that characterization. Anatomically, both pain and temperature receptors are seen as free nerve endings under the microscope.
Dermatomes
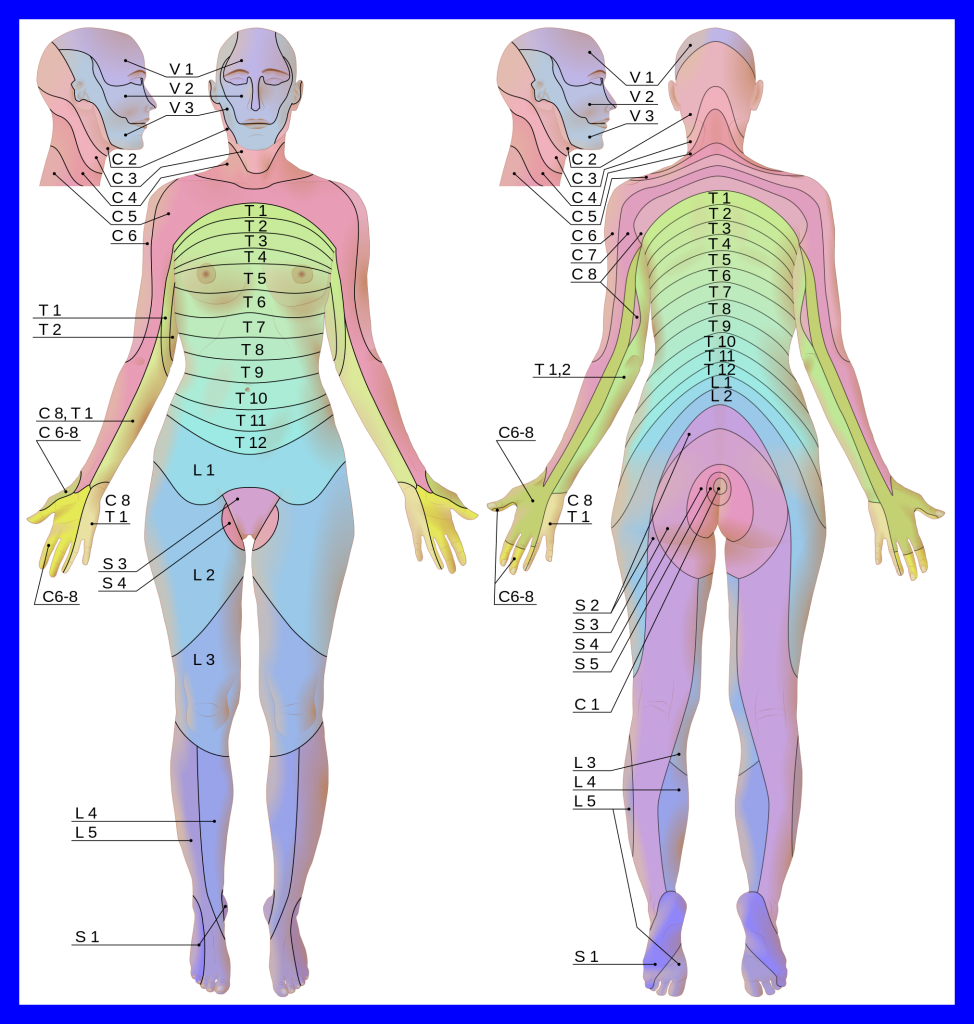
Because a single spinal nerve carries information from a defined region of the skin surface to a single dorsal (posterior) root in the spinal cord we can pair regions of the skin surface with dorsal roots. A map of this pairing is called a dermatomal map and the skin slices on the map are called dermatomes (Greek δερμα, derma–, “skin” + ηομος, –tomos, “slice”).
When a clinician uses a pin to test sensation on various parts of the skin surface, they are mapping dermatomes to see if damage can be located. The varicella virus (a herpesvirus, causing herpes zoster) often lies dormant in a single dorsal root ganglion for most of a patient’s lifetime and then busts out later, infecting the skin surface in a single dermatome, a painful condition called shingles. This can be prevented with the cleverly-named vaccines Zostavax™ or Shingrex™; now you see where these names come from.
We don’t need to learn all the dermatomes, and there’s some variability in the map between different individuals anyway, but a few “landmark” dermatomes are handy and you should learn them:
- C6 and C7 for the thumb and index finger (“C-six shooter”)
- T4 for the nipple line
- T10 for the umbilicus (“belly button”: see the shape of the zero)
- L1-L5 for the Lower extremities.
The dermatome map is evident because a group of unipolar neurons from a patch of skin “real estate” travels together into the central nervous system as part of a spinal nerve. Spinal nerves, somewhat confusingly, change their naming pattern between cervical and lower levels of the vertebral column. The C1 nerve arises from above the C1 vertebra (atlas); the C2 nerve from above the C2 vertebra (axis); and so forth.
The C7 nerve then arises from above the C7 vertebra, the most inferior of the cervical vertebrae. But then there is a C8 nerve that arises from underneath the C7 vertebra.
For the remainder of the spinal nerves, the named nerve arises below the named vertebra. So the T1 spinal nerve arises from below T1, the T2 spinal nerve from below T2, and so forth, for the remainder of the spinal nerves.
Dorsal Columns (Posterior Columns)
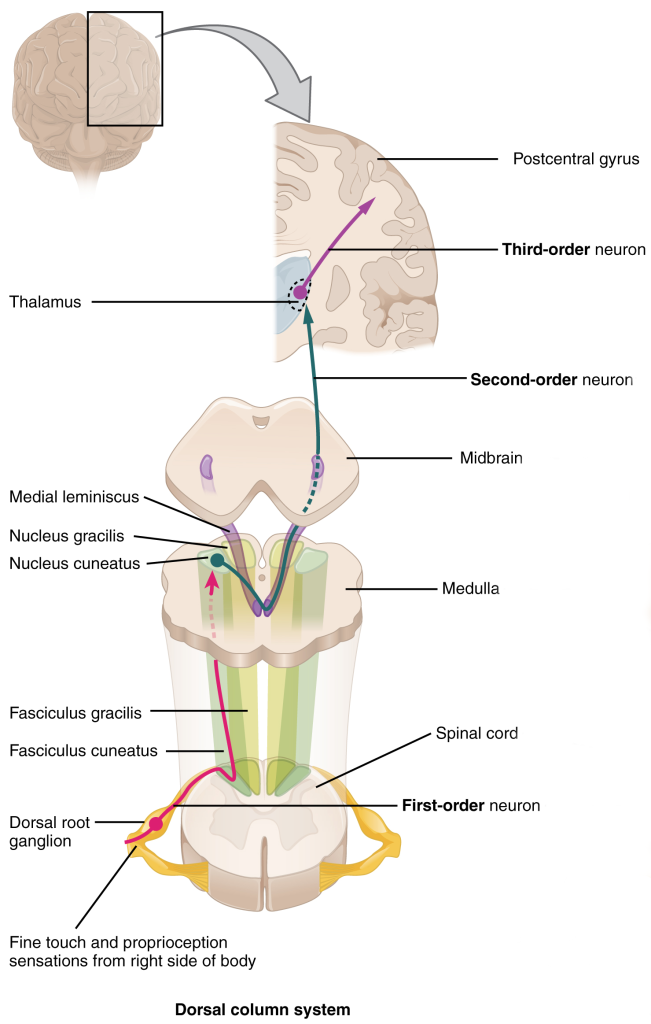
The posterior columns (dorsal columns) carry information about light touch, proprioception, and vibration. As the name tells us, they are found in the posterior (dorsal) part of the spinal cord. Light touch and vibration are probably self-explanatory, but the other two modalities may not be.
Proprioception is the sense of knowing where your body is in space. It involves integrating information from your vestibular (balance) system; muscle stretch; tendon stretch; and skin deflection into a single image of body position. We will talk about each of these sensations separately throughout this Unit.
The posterior columns are comprised of the fasciculus gracilis (“thin bundle of sticks”), carrying information from the lower body, below T4; and the fasciculus cuneatus (“wedge-shaped bundle of sticks”), carrying information from the upper body, above T4.
The posterior columns carry information that needs to be conveyed rapidly to the brain, so it does not stop to talk to other neurons, but rather stays on the same side of the spinal cord as where it entered (i.e. is ipsilateral) and then goes directly to the medulla, the lowest part of the brainstem. There, it synapses in the nuclei of the same name (i.e., the fasciculus gracilis terminates in the nucleus gracilis; the fasciculus cuneatus terminates in the nucleus cuneatus).
The axons leaving the nuclei gracilis and cuneatus then cross as the so-called sensory decussation, or more formally, internal arcuate fibers. They then gather together, now on the opposite side from where the information entered, and are called the medial lemniscus. For this reason, the entire pathway described here is called the posterior column/medial lemniscal system (PCMLS) or dorsal column/medial lemniscal system (DCMLS).
The medial lemniscus ends in the thalamus, contralateral to where the information originated. (We don’t need to know it, but the exact name for this location is the ventroposterolateral or VPL thalamus.)
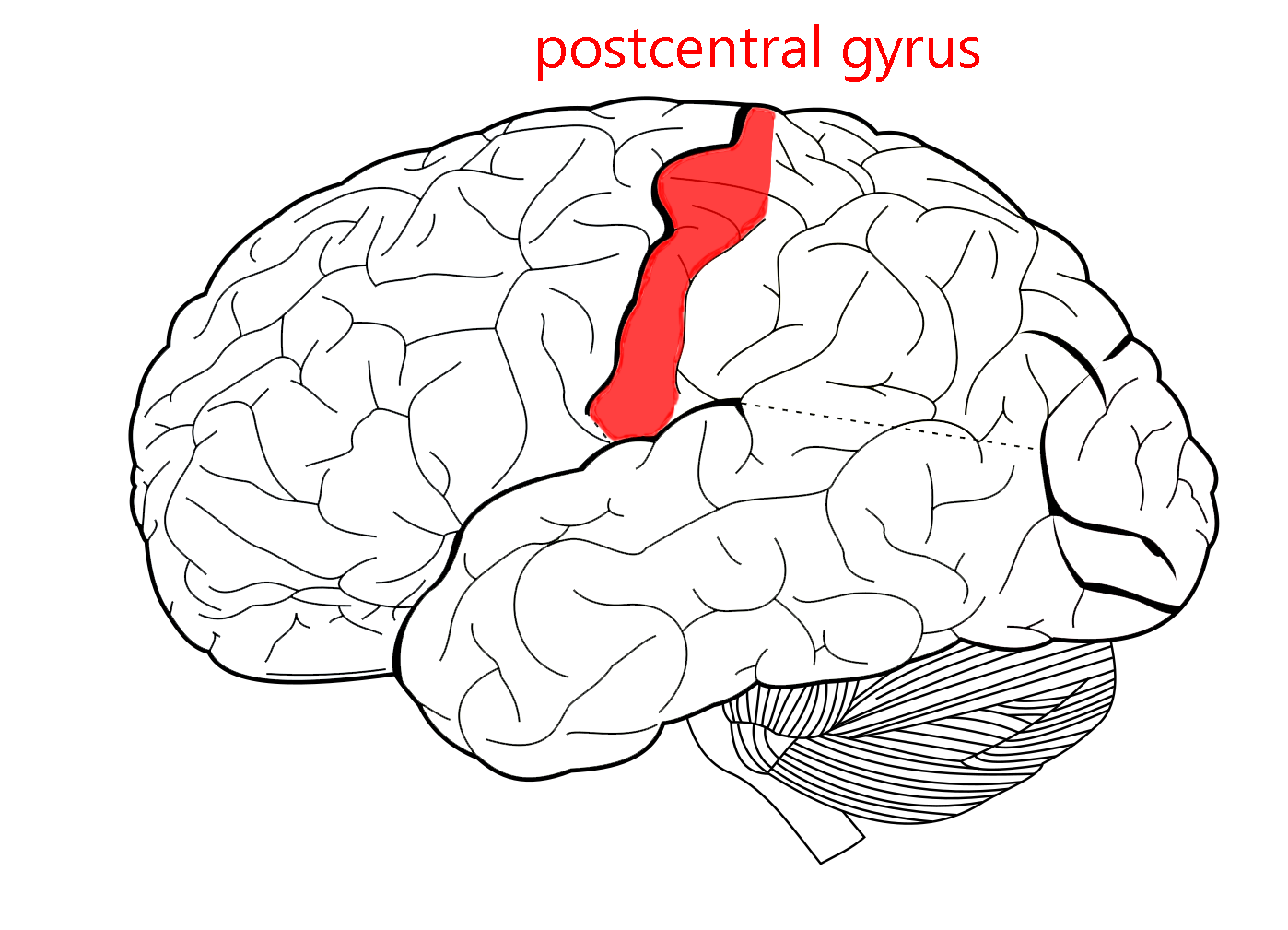
Axons leaving the thalamus then travel through the posterior limb of the internal capsule and corona radiata to the primary somatosensory cortex, anatomically the postcentral gyrus.
Anterolateral System
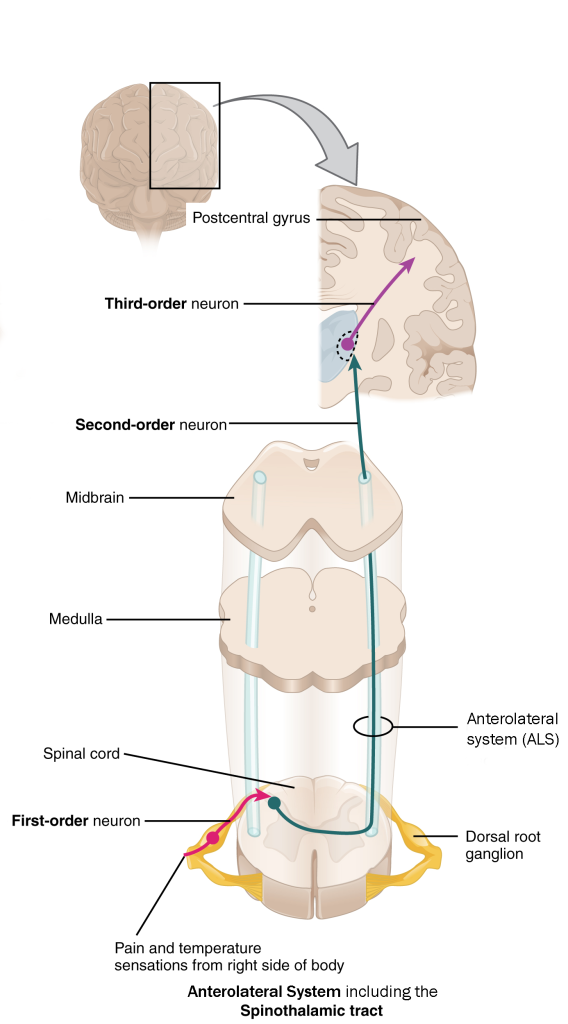
The anterolateral system (ALS) carries information from the spinal cord to the medulla, midbrain, and thalamus. The portion of the ALS that innervates the thalamus is sometimes called the spinothalamic tract. The ALS carries pain and temperature information.
Pain and temperature information is picked up by free nerve endings in the skin, and is carried on spinal nerves and enters on dorsal roots. Inside the spinal cord, it is processed through one or more interneurons before a relay cell axon carries it in the contralateral (opposite side) anterolateral system. Most of the axons terminate in the thalamus.
A minority of the axons innervate nuclei in the medulla and midbrain (raphé and periaqueductal gray). These nuclei, in turn, send axons back to the posterior horn of the spinal cord where they release chemical signals that modify pain perception.
The processing of pain and temperature information in the posterior horn gray matter of spinal cord allows the brainstem and hypothalamus to heavily influence the perception of pain through axons from the brain that interact with the incoming pain signal. Understanding this is of tremendous importance for pain management in patients.
Location of Posterior Columns and Anterolateral System in a Cross-Section of the Spinal Cord
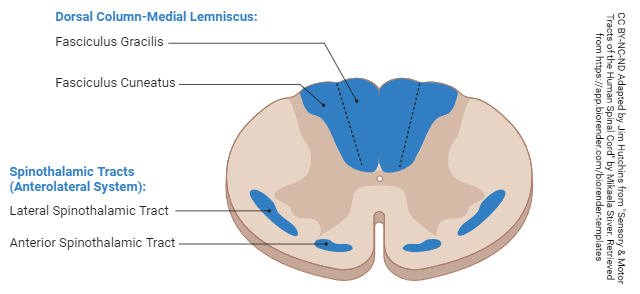
The location of the posterior columns (dorsal columns) and the anterolateral system (old name: spinothalamic tract) is shown in the above diagram.
Somatosensory Cortex
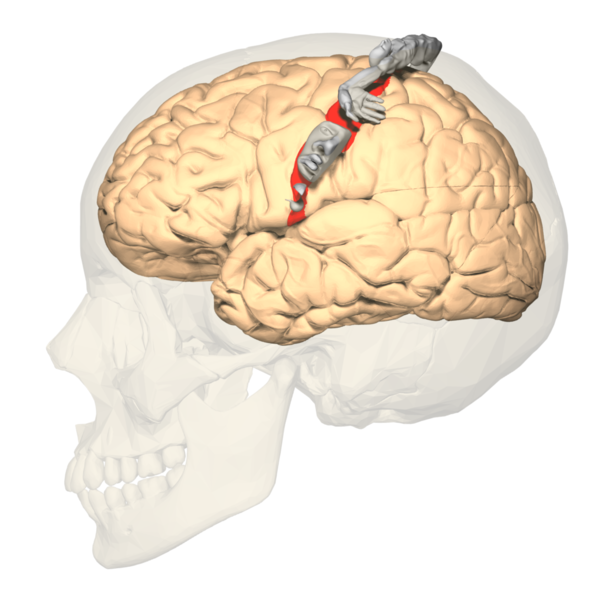
The somatosensory cortex is where both the posterior column—medial lemniscus pathway and anterolateral system pathway terminate.
In order for information from the skin to reach conscious perception, it has to be relayed through the thalamus. Eventually, a neuron carries the skin sensation information to the cortex.
The primary somatosensory cortex is found in Brodmann’s areas 3, 1 and 2 (they are usually listed in this order, no one knows exactly why). This is also the postcentral gyrus of the parietal lobe of the brain, just posterior to the central sulcus.
Earlier, we learned that the precentral gyrus was the same as primary motor cortex and the postcentral gyrus is the same as primary somatosensory cortex.
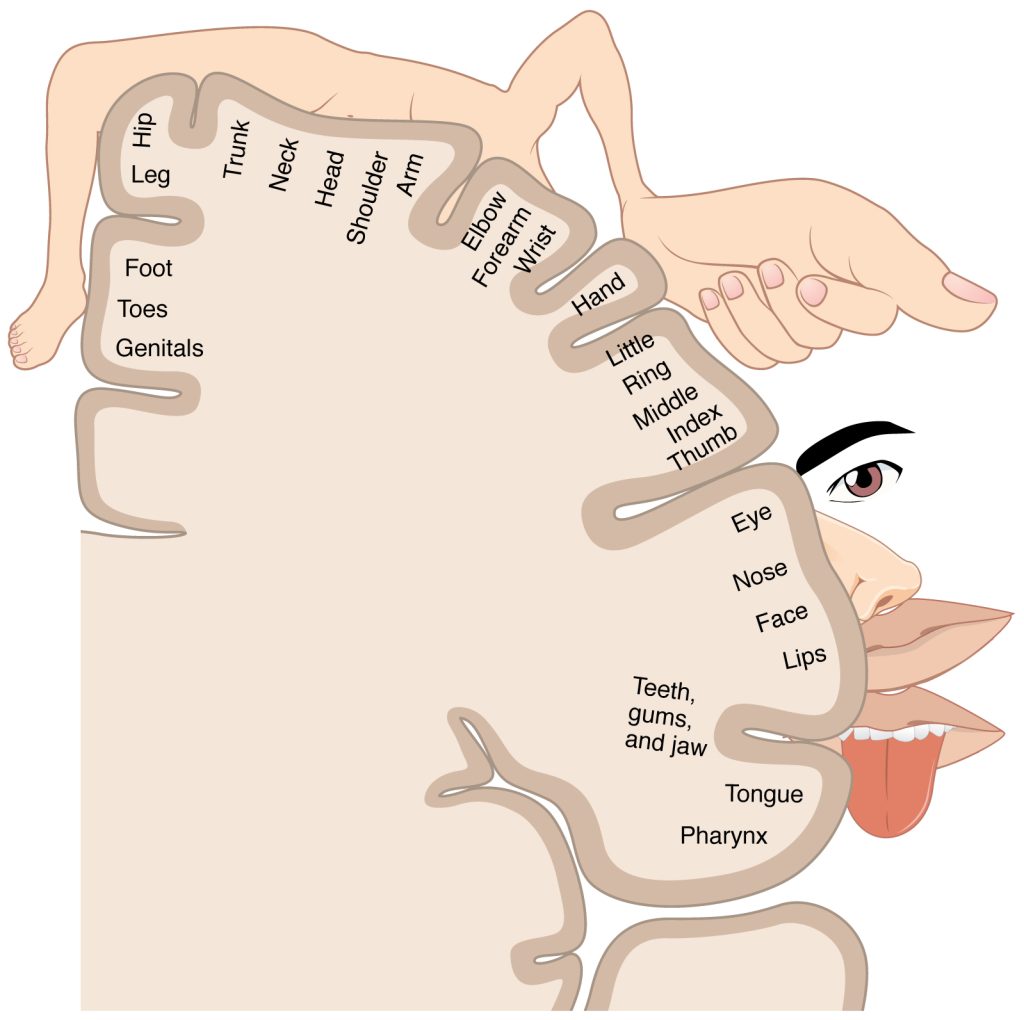
 There is a regular map of the body surface on each of these gyri, as shown here. For the somatosensory cortex, as we travel from lateral to medial, we encounter the gut tube, mouth, face, fingers, hand, arm, back of head, neck, trunk, hip, leg, foot, toes and genitals. Additionally, these maps show the amount of brain “real estate” devoted to each region of the body surface: there is a lot of sensory information coming from lips and hands, since these areas are quite sensitive, and a lot of motor information going out to face and hands, since we have fine control over these areas. This map is called a homunculus (“little man”).
There is a regular map of the body surface on each of these gyri, as shown here. For the somatosensory cortex, as we travel from lateral to medial, we encounter the gut tube, mouth, face, fingers, hand, arm, back of head, neck, trunk, hip, leg, foot, toes and genitals. Additionally, these maps show the amount of brain “real estate” devoted to each region of the body surface: there is a lot of sensory information coming from lips and hands, since these areas are quite sensitive, and a lot of motor information going out to face and hands, since we have fine control over these areas. This map is called a homunculus (“little man”).
Media Attributions
- U12-034 Skin Layers © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license
- U12-034a Sensory Receptors of Skin © Molnar, Charles and Gair, Jane is licensed under a CC BY (Attribution) license
- U12-035 Meissner Corpuscles © Wbensmith is licensed under a CC BY (Attribution) license
- U12-036 Pacinian Corpuscles © Uthman, Ed is licensed under a CC BY (Attribution) license
- U12-037 Skin Free Nerve Endings © BruceBlaus is licensed under a CC BY (Attribution) license
- U12-038 Somatosensation Sensory Input © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license
- U12-033 Mechanically-gated Channel © Cornelius, Emma is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- U12-039 Somatosensory Receptors with locations © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter adapted by Bizzell, Lizz is licensed under a CC BY (Attribution) license
- U12-040 Dermatomes Labeled Female Front Back © Goran_tek-en is licensed under a CC BY-SA (Attribution ShareAlike) license
- U12-041 Somatosensory Pathways Dorsal Columns © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- U12-042 Somatosensory Pathways ALS © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter adapted by Jim Hutchins is licensed under a CC BY (Attribution) license
- U12-043a Sensory Tracts of the Human Spinal Cord v2 © Hutchins, Jim and Stiver, Mikaela is licensed under a CC BY-NC-ND (Attribution NonCommercial NoDerivatives) license
- U11-072 Postcentral Gyrus © Carter, Henry Vandyke adapted by Jim Hutchins is licensed under a CC BY-SA (Attribution ShareAlike) license
- U12-044 Primary Somatosensory Cortex Lateral View © Unknown is licensed under a CC BY-SA (Attribution ShareAlike) license
- U12-045 Sensory Homunculus © Betts, J. Gordon; Young, Kelly A.; Wise, James A.; Johnson, Eddie; Poe, Brandon; Kruse, Dean H. Korol, Oksana; Johnson, Jody E.; Womble, Mark & DeSaix, Peter is licensed under a CC BY (Attribution) license

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}